
Production
and
characterization
of
exocellular
proteases
in
ectomycorrhizal
fungi
K.
El-Badaoui
B. Botton
Universitc
de
Nancy
I,
Facult6
des
Sciences,
Laboratoire
de
Physiologie
V6g6tale
et
Foresti6re,
BP
239,
54506
Vandœuvre-Iès-Nancy
Cedex,
France
Introduction
Mycorrhizas
grow
very
often
in
soil
layers
in
which
organic
nitrogen
compounds
are
present
in
large
quantities.
The
ability
of
several
ectomycorrhizal
fungi
to
assimilate
proteins
and
to
transfer
their
nitrogen
to
plants
has
already
been
demonstrated
(Abuzinadah
and
Read,
1986;
Abuzinadah
et
al.,
1986).
These
results
suggest
that
proteases
are
excreted
by
the
fungi
and
protease
activities
detected
in
mycorrhiza-
forming
fungi
were
reported
by
Lyr
(1963)
and
Ramstedt
and
S6derhall
(1983).
In
this
study,
the
process
of
production
of
exocellular
proteases
has
been
investi-
gated
in
free
living
fungi
as
well
as
in
mycorrhizas.
Experiments
were
carried
out
with
Amanita
rubescens
and
Lacta-
rius
subdulcis
which
live
in
organic
hori-
zons
and
with
Cenococcum
geophilum
and
Hebeloma
crustuliniforme
which
live
predominantly
in
mineral
soil
layers.
Materials
and
Methods
The
fungi
were
collected
from
mycorrhizas.
They
were
grown
in
shaken
cultures
at
25°C
in
Pachlewski’s
medium
containing
nitrogen
as
either
diammonium
tartrate
or
proteins:
bovine
serum
albumin
(BSA),
gelatin,
casein
or
pro-
teins
extracted
from
the
litter
(Botton
et
al.,
1986).
Enzyme
activities
were
determined
in
culture
filtrates,
either
by
spectrophotometry
or
by
spectrofluorometry
using
fluorescein
isothio-
cyanate-labeled
BSA
(FITC-BSA)
as
described
by
Twining
(1984).
Proteins
purified
from
the
litter
by
ammonium
sulfate
fractionation
and
DEAE-cellulose
chromatography
were
also
labeled
with
FIT’C
and
used
as
a
substrate
and
as
inducers.
Results
Protease
activity
remained
at
a
very
low
level
when
the
fungus
was
cultivated
in
the
presence
of
ammonium,
but
increased
strongly
after
transfer
of
the
organism
to
fresh
medium
containing
casein,
gelatin
or
BSA
as
the
sole
nitrogen
source.
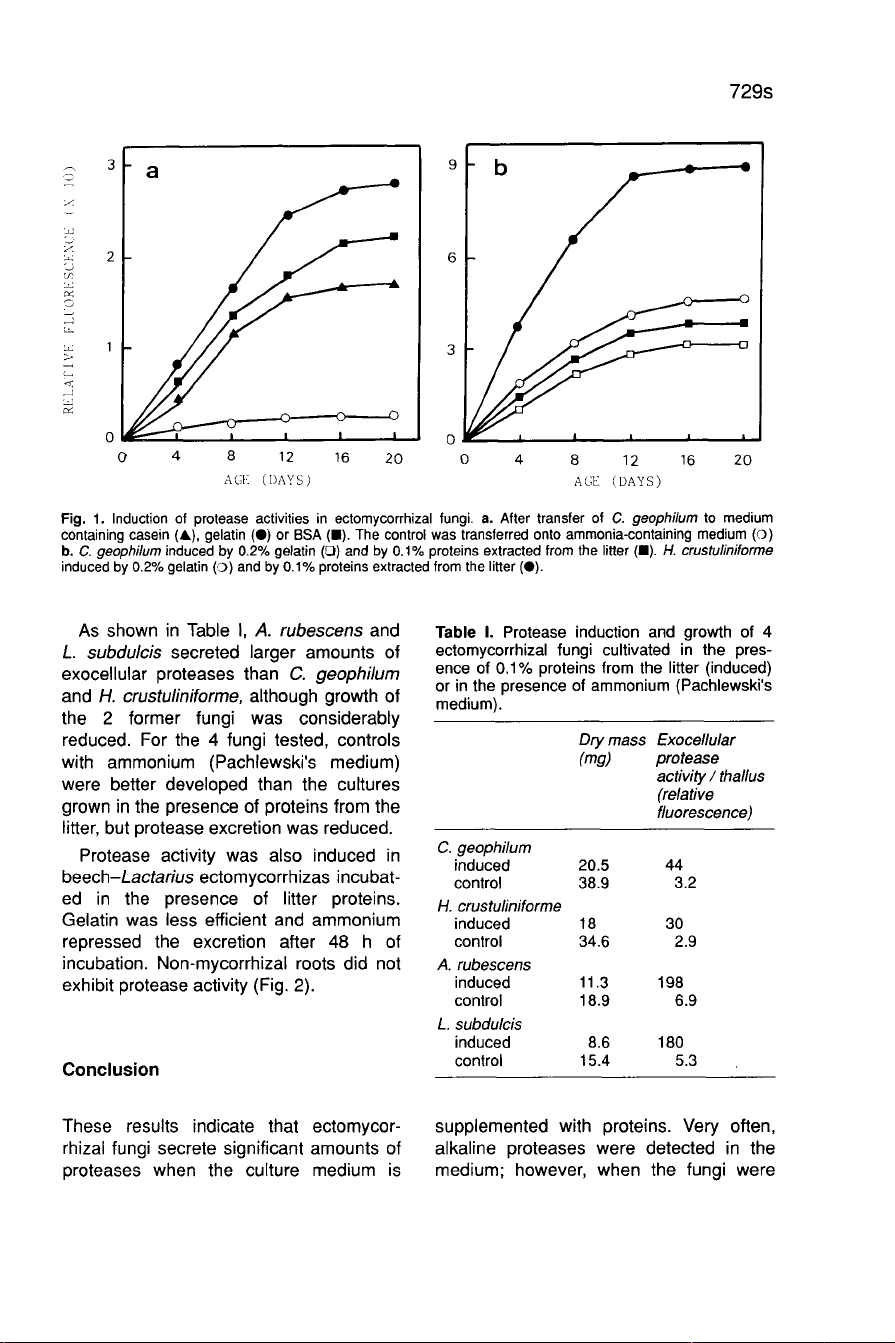
Gelatin
proved
to
be
the
most
efficient
inducer
(Fig.
1a).
Elimination
of
carbon,
inorganic
nitrogen
or
sulfur
was
not
sufficient
to
induce
exoceillular
proteases,
proteins
also
had
to
be
present.
However,
in
nitrogen-
and
carbon-deficient
media,
gelatin
was
not
efficient
(not
shown).
A
group
of
proteins
of
about
46
kDa
was
purified
from
the
forest
litter
and
was
a
better
inducer
than
gelatin,
in
spite
of
its
lower
concentration,
both
in
C.
geophilum
and
H.
crustuliniforme
(Fig.
1 b).
As
shown
in
Table
I,
A.
rubescens
and
L.
subdulcis
secreted
larger
amounts
of
exocellular
proteases
than
C.
geophilum
and
H.
crustuliniforme,
although
growth
of
the
2
former
fungi
was
considerably
reduced.
For
the
4
fungi
tested,
controls
with
ammonium
(Pachlewski’s
medium)
were
better
developed
than
the
cultures
grown
in
the
presence
of
proteins
from
the
litter,
but
protease
excretion
was
reduced.
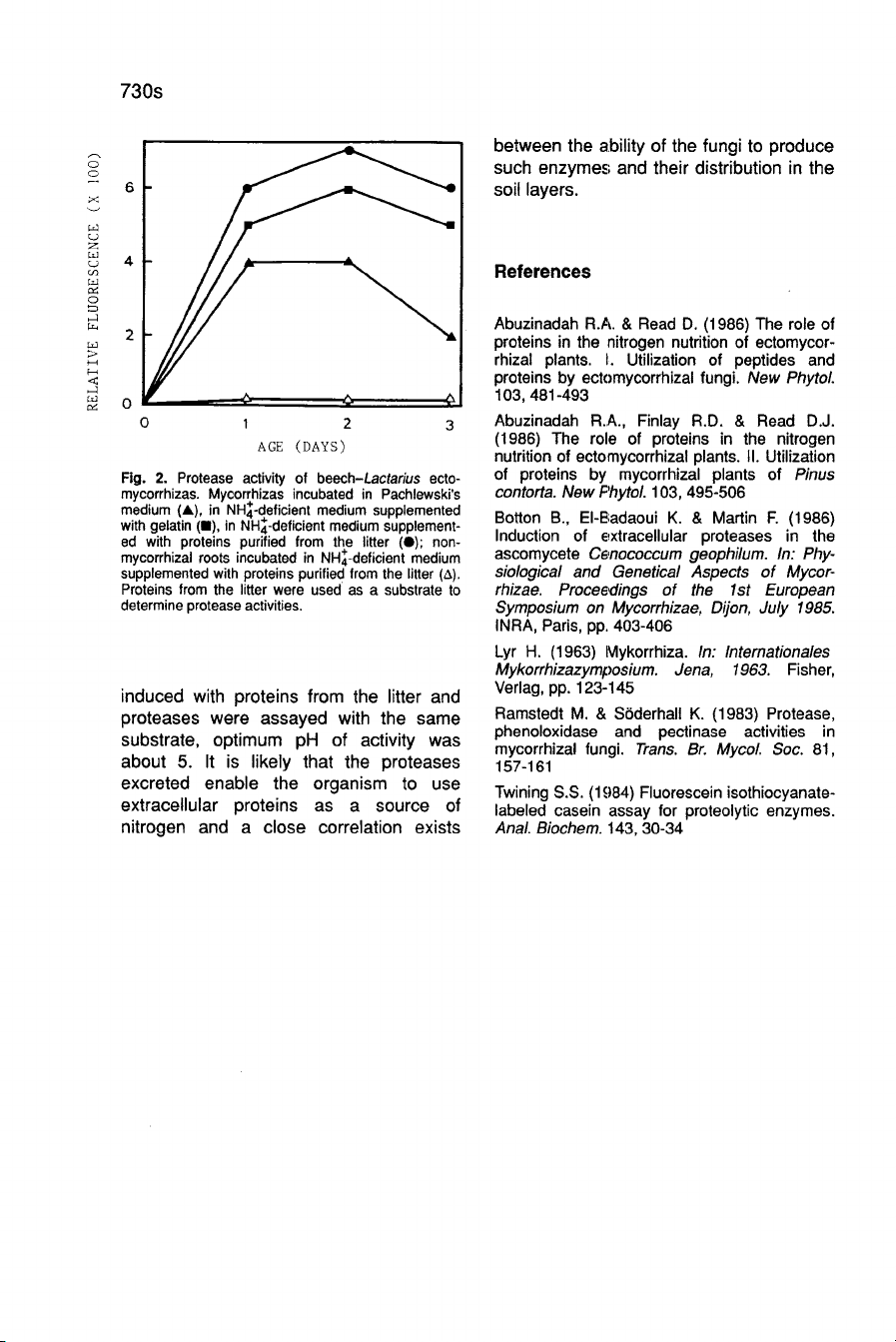
Protease
activity
was
also
induced
in
beech-Lactarius
ectomycorrhizas
incubat-
ed
in
the
presence
of
litter
proteins.
Gelatin
was
less
efficient
and
ammonium
repressed
the
excretion
after
48
h
of
incubation.
Non-mycorrhizal
roots
did
not
exhibit
protease
activity
(Fig.
2).
Conclusion
These
results
indicate
that
ectomycor-
rhizal
fungi
secrete
significant
amounts
of
proteases
when
the
culture
medium
is
supplemented
with
proteins.
Very
often,
alkaline
proteases
were
detected
in
the
medium;
however,
when
the
fungi
were
induced
with
proteins
from
the
litter
and
proteases
were
assayed
with
the
same
substrate,
optimum
pH
of
activity
was
about
5.
It
is
likely
that
the
proteases
excreted
enable
the
organism
to
use
extracellular
proteins
as
a
source
of
nitrogen
and
a
close
correlation
exists
between
the
ability
of
the
fungi
to
produce
such
enzymes
and
their
distribution
in
the
soil layers.
References
Abuzinadah
R.A.
&
Read
D.
(1986)
The
role
of
proteins
in
the
nitrogen
nutrition
of
ectomycor-
rhizal
plants.
I.
Utilization
of
peptides
and
proteins
by
ectomycorrhizal
fungi.
New
Phytol.
103, 481-493
Abuzinadah
R.,A.,
Finlay
R.D.
&
Read
D.J.
(1986)
The
role of
proteins
in
the
nitrogen
nutrition
of
ectomycorrhizal
plants.
11.
Utilization
of
proteins
by
mycorrhizal
plants
of
Pinus
contorta.
New F’hytoG
103,
495-506
Botton
B.,
El-Badaoul
K.
&
Martin
F.
(1986)
Induction
of
extracellular
proteases
in
the
ascomycete
Cenococcum
geophilum.
In:
Phy-
siological
and
Genetical
Aspects
of
Mycor-
rhizae.
Proceedings
of
the
Ist
European
Symposium
on
Mycorrhizae,
Dijon,
July
1985.
INRA,
Paris,
pp.
403-406
Lyr
H.
(1963)
IVlykorrhiza.
In:
Infernationales
Mykorrhizazymposium.
Jena,
1963.
Fisher,
Verlag,
pp.
123-145
Ramstedt
M.
&
S6derhall
K.
(1983)
Protease,
phenoloxidase
and
pectinase
activities
in
mycorrhizal
fungi.
Trans.
Br.
Mycol.
Soc.
81,
157-161
Twining
S.S.
(1984)
Fluorescein
isothiocyanate-
labeled
casein
assay
for
proteolytic
enzymes.
Anal.
Biochem.
143, 30-34
















%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)