
Original
article
Cytogenetic
study
of
diploid
and
spontaneous
triploid
oaks,
Quercus
robur
L
AK
Butorina
Department
of
Genetics
and
Bioecology
Central
Research
Institute
of
Forest
Genetics
and
Breeding,
Voronezh,
Russia
Summary —
Data
are
presented
on
the
cytogenetics
of
2
unusually
large
oak
trees
found
in
the
Voronezh
region
of
Russia.
In
both
trees,
cells
with
2n
=
3x
=
36
chromosomes
were
predominant,
with
occasional
observations
of
diploid
hypoaneuploid
and
hyperaneuploid
cells.
Functionally,
the
trees
can
be
considered
triploids,
although
in
a
strict
sense,
they
are
mixoploids.
Meiosis
in
micro-
sporogenesis
of
these
trees
is
very
disturbed
and,
as
a
consequence,
pollen
with
unbalanced
chro-
mosome
numbers
are
produced.
Correspondingly,
the
progeny
from
each
tree
were
very
different
in
morphological
characteristics
and
cytogenetic
constitution.
These
progeny
can
be
used
in
gene
mapping
studies
and
for
other
basic
research
purposes.
Studies
on
some
diploid
oaks
reveal
the
presence
of
2n
pollen,
formed
by
parallel
spindles
in
the
2nd
meiotic
division.
Methods
for
producing
additional
oak
triploids
that
have
a
potential
for
heterosis
are
discussed.
oak
/
triploid
/
mixoploid
/
meiosis
/
meiotic
mutant
/
progeny
Résumé —
Cytogénétique
de
chênes
diploïdes
et
triploïdes
spontanés
(Quercus
robur
L).
Des
données
cytogénétiques
relatives
à
2
chênes
de
très
grande
taille
situés
dans
la
région
de
Vo-
ronezh
(Russie)
sont
présentées
dans
cet
article.
Des
cellules
comprenant
2n
=
3x
=
36
chromo-
somes
sont
prédominantes
dans
chaque
arbre.
Des
cellules
hypoaneuploïdes
et
hyperaneuploïdes
ont
également
été
rencontrées.
Au
plan
fonctionnel,
les
2
arbres
peuvent
être
considérés
comme
tri-
ploïdes,
plus
précisément
mixoploïdes.
La
méiose
durant
la
microsporogenèse
est
très
perturbée
et
produit
des
grains
de
pollen
au
nombre
de
chromosomes
déséquilibré.
Les
descendants
de
ces
arbres
manifestent
des
caractéristiques
morphologiques
et
une
constitution
cytogénétique
très
va-
riables.
Ces
familles
peuvent
être
utilisées
pour
des
études
de
cartographie
génétique
et
d’autres
re-
cherches
fondamentales.
Des
études
similaires
faites
sur
des
chênes
diploïdes
mettent
en
évidence
des
grains
de
pollen
à
2n
chromosomes
formés
par
des
fuseaux
parallèles
lors
de
la
seconde
divi-
sion
méiotique.
Des
méthodes
de
production
de
chênes
triploïdes
en
vue
de
générer
de
l’hétérosis
sont
discutées.
chêne
/ triploïde
/
mixoploïde
/ méiose
/ mutant
méiotique
/ descendance
INTRODUCTION
The
Voronezh
region
in
south
central
Rus-
sia
is
famous
for
oak
stands
producing
ex-
cellent
quality
lumber.
Rich
Voronezh
chernozem
provide
optimum
edaphic
conditions
for
oak
species.
Two
triploid
trees
of
Quercus
robur
L
were
discovered
in
the
Voronezh
region
by
the
scientific
re-
searchers
of
the
Central
Research
Insti-
tute
of
Forest
Genetics
and
Breeding,
VV
levlev
and
TI
Pletmintseva.
The
trees
were
more
than
100
years
old
and
differed
from
oaks
of
a
similar
age.
They
were
of
gigan-
tic
height,
weak
fertility
and
exhibited
unusual
morphological
and
anatomical
features.
These
characteristics
have
often
indicated
a
polyploid
nature
in
many
plant
species.
In
order
to
obtain
objective
information
about
the
cytogenetic
nature
of
these
2
trees,
we
analyzed
various
cytological
characteristics,
including
chromosome
number,
meiosis
in
microsporogenesis
and
the
development
of
the
male
gameto-
phyte.
MATERIALS
AND
METHODS
For
the
cytological
investigations,
branches
were
taken
from
each
putative
triploid
and
a
number
of
putative
diploid
trees
at
the
appropriate
sampling
period
for
meiotic
observations.
The
branches
were
placed
into
water
vessels
and
kept
in
a
cold
room.
Somatic
chromosome
counts
were
made
using
vegetative
buds
that
were
removed
from
the
branches
and
placed
in
a
damp
penicillin
bot-
tle
under
freezing
conditions
for
1-2
h
to
inhibit
spindle
fiber
formation.
The
young
leaves
were
then
fixed
in
aceto
alcohol.
For
the
study
of
meiosis
in
microsporogene-
sis
and
the
process
of
development
of
the
male
gametotype,
the flower
buds
were
fixed
from
the
stage
of
green
cone
up
to
the
flowering.
All
materials
were
stained
in
acetohaemotac-
silin.
The
squash
technique
was
used
to
prepare
slides
for
microscopical
examination.
RESULTS
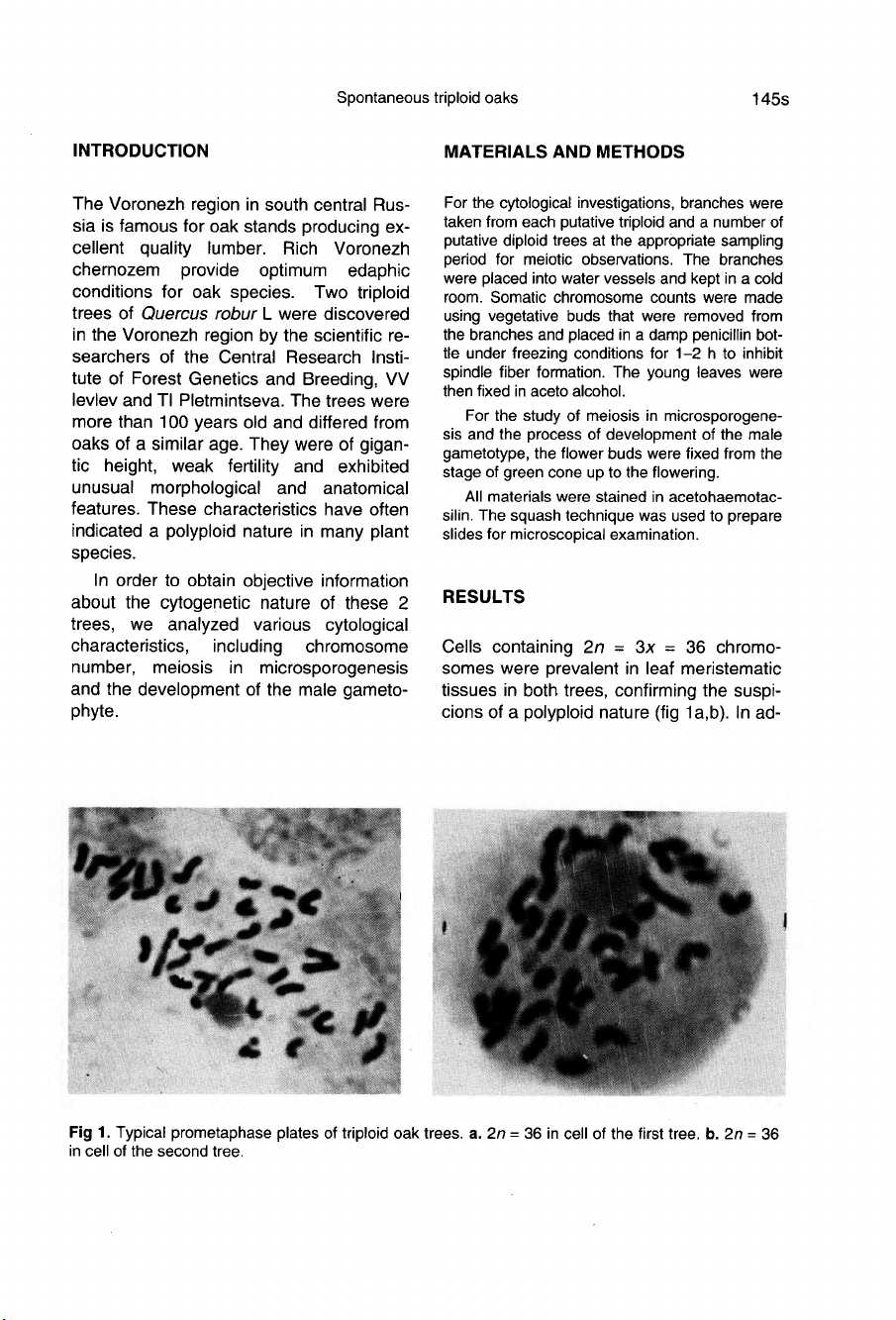
Cells
containing
2n
=
3x
=
36
chromo-
somes
were
prevalent
in
leaf
meristematic
tissues
in
both
trees,
confirming
the
suspi-
cions
of
a
polyploid
nature
(fig
1a,b).
In
ad-

dition
to
the
triploid
cells,
each
tree
had
meristematic
cells
with
other
chromosome
numbers.
Diploid,
hypoaneuploid
and
hy-
peraneuploids
cells
were
found.
Conse-
quently,
these
trees
could
be
classified
as
mixoploids
in
a
strict
sense.
Meiosis
in
both
trees
was
abnormal,
as
typically
found
in
plants
with
unbalanced
chromosome
numbers.
At
metaphase
I,
a
broad
spectrum
of
chromosomal
configura-
tions,
from
36
univalents
to
12
trivalents,
were
observed.
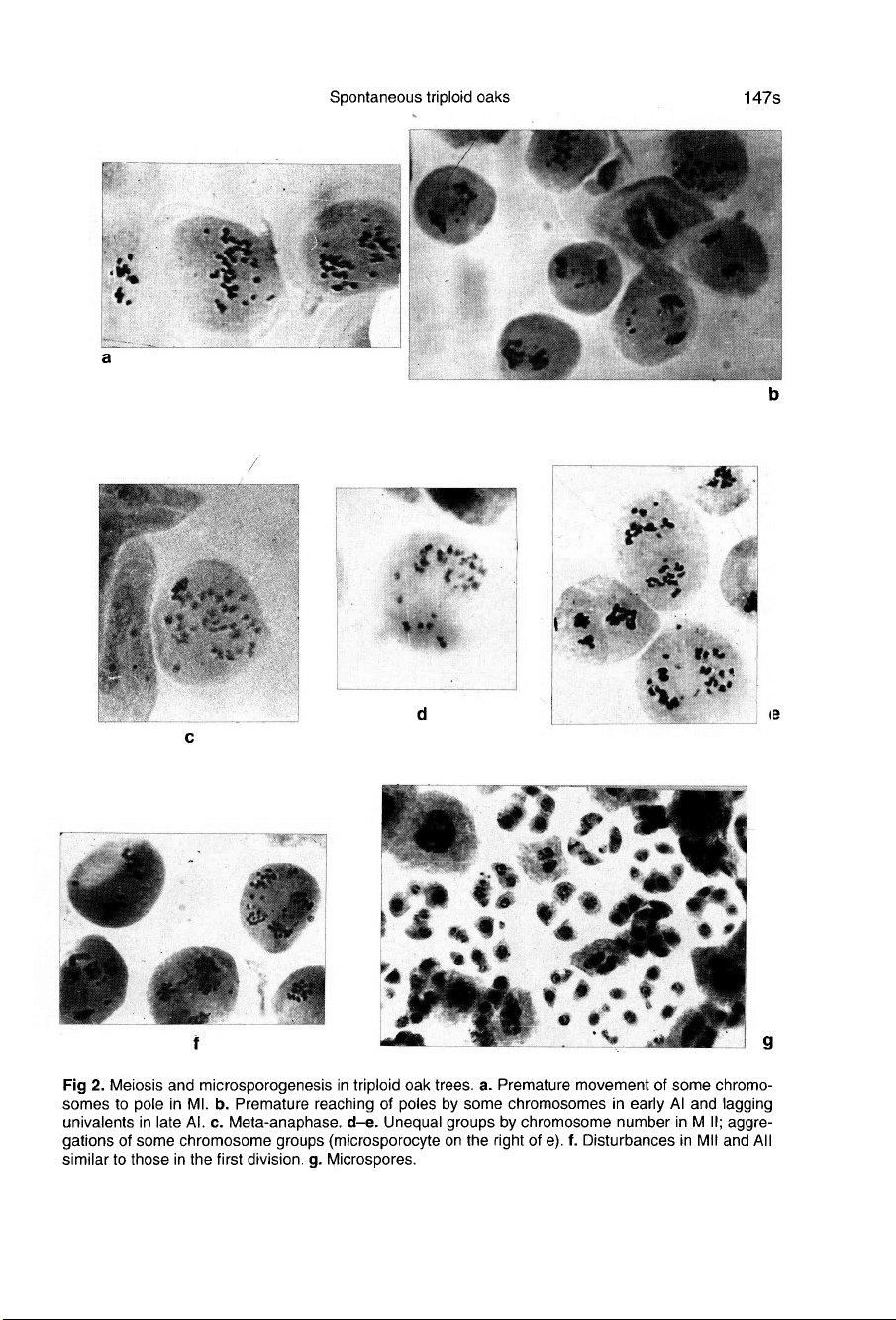
Lagging
chromosomes
were
frequent
in
anaphase
I (fig
2a,b).
Some
chromosomes
were
delayed
at
the
equatorial
plate,
while
others
were
located
outside
the
achromatic
spindle
(fig
2b).
Oc-
casionally,
metaphase
plates
were
formed
and
only
one
stage
was
observed:
meta-
anaphase
(fig
2c).
In
such
instances,
the
distribution
of
chromosomes
in
metaphase
II
was
unequal
(fig
2d,e).
In
some
micro-
sporocytes,
aggregation
of
chromosomes
into
separate
groups
was
observed
(fig
2e).
The
chromosomal
disturbances
in
the
second
meiotic
division
were
similar
to
ab-
normalities
in
the
first
division
(fig
2f).
Cor-
respondingly,
many
unbalanced
micro-
spores
were
formed
(fig
2g),
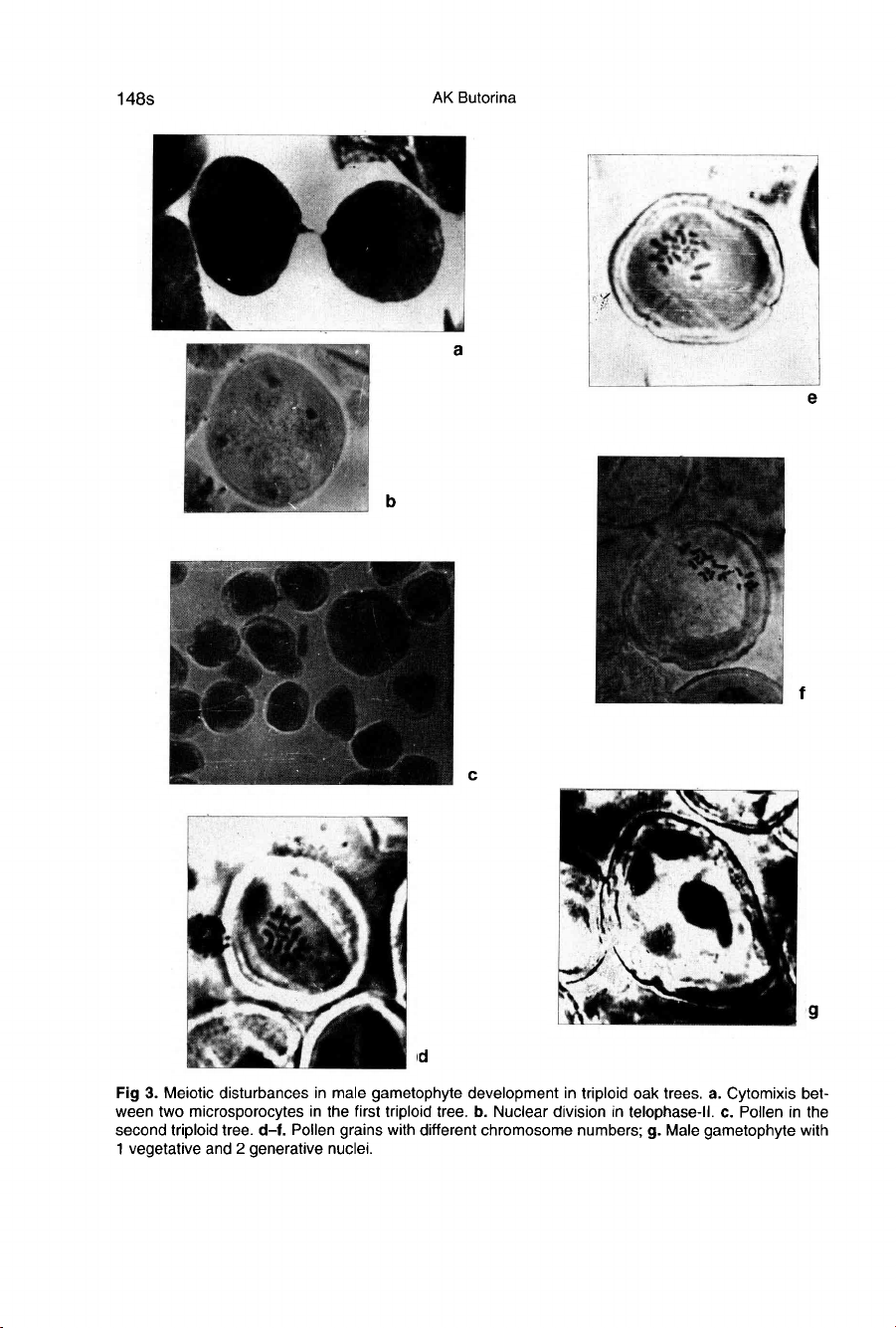
that
subsequently
resulted
in
pollen
grains
with
different
chromosome
numbers
(fig
3a-c).
The number
of
cells
with
meiotic
distur-
bances
in
both
oak
trees
varied
over
differ-
ent
years.
The
maximum
percentage
of
ab-
normal
divisions
was
98%
of
the
total
number
of
dividing
microsporocytes.
Although
the
2
triploid
oak
trees
had
many
common
meiotic
characteristics,
they
also
had
specific
peculiarities.
Cyto-
mixis
was
observed
in
one
tree
(fig
3a).
Preliminary
divisions
of
the
nuclei
in
telo-
phase
II
were
found
in
the
other
tree,
re-
sulting
in
unbalanced
pollen
(fig
3b).
The
meiotic
irregularities
caused
diversity
in
pollen
chromosome
number
and
distur-
bances
in
male
gametophyte development
(fig
3d-g).
The
same
anomalies
may
be
ex-
pected
in
the
female
gametophyte,
since
the
progeny
of
these
trees
have
variable
morphological
characters
(levlev
and
Plet-
mintzeva,
personal
communication).
Meiosis
in
pollen
mother
cells
of
diploid
oak
trees
was
also
investigated.
This
pro-
cess
was
found
to
be
essentially
normal
in
these
trees,
with
approximately
5%
of
the
microsporocytes
exhibiting
abnormal
divi-
sion.
However,
among
these
trees,
several
individuals
were
found
to
form
2n
pollen
grains,
comprising
5-10%
of
the micro-
spores
(fig
4a).
The
2n
pollen
was
found
to
be
formed
by
parallel
spindles
(sensu,
Mok
and
Peloquin,
1975)
in
the
second
meiotic
division
(fig
4b).
DISCUSSION
Chromosomal
variation
in
progeny
from
the
triploid
trees
can
be
the
basis
for
fur-
ther
cytogenetic
research.
In
particular,
analyses
of
aneuploid
offspring
from
the
triploid
trees
would
be
an
excellent
method
to
genetically
map
oak
chromosomes.
Un-
fortunately,
only
these
2
triploid
oaks
are
known
to
exist.
Other
mature
triploids
of
oak
have
not
been
discovered.
A
possible
cause
may
relate
to
dysgenic
selection
that
was
conducted
in
forests
of
this
region
for
many
years.
It
would
be
desirable
to
study
additional
trees
in
order
to
gain
a
better
understanding
of
the
mechanism(s)
of
origin
for
triploid
oaks.
Other
triploid
Q
robur
have been
ob-
served
in
studies
of
twin
seedlings
by
Johnsson
(1946)
and
Burda
and
Schepo-
tiev
(1973).
These
scientists
respectively
speculated
that
polyembryony
could
be
re-
sponsible
for
the
triploid
condition.
Unfortu-
nately,
there
have
been
no
additional
re-
ports
on
the
triploid
seedlings
identified
in
their
studies.
Our
data
suggests
that
trip-
loid
oaks
may
originate
by
participation
of


























%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)