
Original article
Lack of mother tree alleles in zymograms
of Cupressus dupreziana A. Camus embryos
Christian Pichot*, Bruno Fady and Isabelle Hochu
INRA, Unité de Recherches Forestières Méditerranéennes, Équipe de Génétique, Av. Vivaldi, 84000 Avignon, France
(Received 12 May 1999; accepted 13 September 1999)
Abstract – Unexpected ploidy levels observed in a previous flow cytometric analysis of Cupressus dupreziana endosperm, led us to
hypothesize an asexual seed production in this endangered species. We tested here this possible apomixis, studying isozyme variabil-
ity of endosperms, embryos and seed coats extracted from open pollinated seeds. Electrophoretic bands observed in zymograms of
PGI2 and LAP, the only two polymorphic systems, were in agreement with bands observed in C. sempervirens zymograms.
Zymograms of C. dupreziana endosperms were identical to those of the diploid maternal tissue, as observed in C. sempervirens.
Hypothesizing a codominant nuclear DNA control for both PGI2 and LAP, a similar electrophoretic expression of these systems in
endosperm and embryo, and the absence of modifier genes, the lack of endosperm bands in most embryo zymograms was interpreted
as the result of a strictly paternal origin of embryo nuclear DNA in C. dupreziana seeds.
Cupressus dupreziana / isozyme / reproduction / gymnosperm / paternal inheritance
Résumé – Absence d’allèles maternels dans les zymogrammes des embryons de Cupressus dupreziana A. Camus. De précé-
dents travaux portant sur les niveaux de ploïdie de l'endosperme des graines de Cupressus dupreziana, nous avaient conduit à émettre
l'hypothèse d'une reproduction apomictique chez cette espèce menacée. Cette hypothèse a été ici testée par l'étude de la variabilité
enzymatique d'endospermes, d'embryons et de téguments de graines issues de pollinisation libre. Les zymogrammes observés pour
les deux systèmes polymorphes, PGI2 et LAP, sont conformes à ceux observés chez C. sempervirens. Les zymogrammes des endo-
spermes de C. dupreziana sont identiques à celui de l'arbre mère, comme précédemment observé chez C. sempervirens. La plupart
des zymogrammes produits par les embryons des graines de C. dupreziana ne contiennent pas les bandes électrophorétiques des
endospermes correspondants. Chez tous les gymnospermes étudiés, PGI2 et LAP sont des marqueurs codominants, sous contrôle
génétique nucléaire. Chez C. sempervirens l’expression de ces systèmes est indépendante des tissus (endosperme ou embryon) et
aucun gène modificateur n’est connu. Dans ce contexte, nous formulons l’hypothèse d’une origine strictement paternelle de l’ADN
nucléaire des embryons.
Cupressus dupreziana / isozyme / reproduction / gymnosperme / hérédité paternelle
1. INTRODUCTION
In gymnosperm seeds, the embryo is generally sur-
rounded by a haploid tissue called endosperm (or
megagametophyte). However we recently demonstrated
that in some Cupressaceous species, and especially in
Mediterranean cypresses, the endosperm exhibits multi-
ple levels of ploidy [33]. Cupressus sempervirens L.
originating from the eastern Mediterranean basin, and
C. atlantica Gaussen, an endemic species from the
Atlas mountain in Morocco, exhibited similar profiles
of endosperm DNA contents, corresponding to even
and odd levels of ploidy (1C, 2C, 3C, 4C, 5C, 6C...).
We demonstrated that these multiple ploidy levels were
Ann. For. Sci. 57 (2000) 17–22 17
© INRA, EDP Sciences 2000
* Correspondence and reprints
Tel. (33) 04 90 13 59 23; Fax. (33) 04 90 13 59 59; e-mail: pichot@avignon.inra.fr
C. Pichot et al.
18
due to nuclei fusion during megagametogenesis [13]. In
gymnosperms, the endosperm derives from the megaga-
metophyte that develops, after meiosis, from generally
one but sometimes several (four in C. sempervirens)
megaspore(s) [14]. Through archegonia, the megagame-
tophyte produces female gamete(s).
Multiple ploidy levels were also observed in the
endosperm of Cupressus dupreziana A. Camus, the other
Mediterranean cypress but, unexpectedly, only even lev-
els [35]. C. dupreziana is an endangered tree species
from the Tassili N’Ajjer desert, Algeria [3, 36].
Approximately 150 trees are still surviving in this natural
area [10]. The absence of 1C DNA nuclei in C.
dupreziana endosperm raises the question of existence
(and origin) of female gametes. In order to test an hypo-
thetical asexual seed production (apomixis), isozyme
variability was studied in 5 open-pollinated families.
2. MATERIALS AND METHODS
Seeds were extracted from mature cones collected
from five 17 year old C. dupreziana trees (referred as
D38, D28, OO1, J1 and DD5) planted in a ex-situ collec-
tion in the Esterel Mountain (Southern France) distant by
1.5 km from the nearest cypress stand which reduces the
possibility of exogenous pollination. This plantation is
composed of 275 C. dupreziana trees representing 35
clones (5 ramets per clone) and 6 open pollinated proge-
nies. Genetic diversity of this material is unknown. Trees
OO1, J1 and DD5 are three different clones, while D38
and D28 belong to the same progeny.
Seeds from one C. sempervirens tree were used as
control. In order to confirm the diploid-like maternal ori-
gin of the endosperm [29], we also analyzed endosperms
and seed coats of immature seeds collected from another
C. dupreziana tree referred as RUS.
Isozyme analyses were performed on samples
(embryo, endosperm or seed-coats) extracted from single
seeds. Isozymes were revealed using horizontal starch
gel electrophoresis. Electrophoresis and staining proce-
dures followed those of Papageorgiou [28]. Four enzyme
systems were scored: Leucine aminopeptidase (LAP, EC
3.4.11.1), Malate dehydrogenase (MDH, EC 1.1.1.37),
Nicotinamide adenine dinucleotide dehydrogenase
(NDH, EC 1.6.99.3) and Phosphoglucose isomerase
(PGI, EC 5.3.1.9). Zones and bands in each system were
coded according to their decreasing mobility, starting
from 1 for the fastest zone or band.
3. RESULTS
Most seeds were empty in all C. dupreziana seed lots
(table I). One seed, from tree DD5, had an endosperm
but no embryo.
No variation was observed in MDH and NDH.
Banding patterns were similar to those of the C. semper-
virens control. PGI and LAP were polymorphic and thus
usable for analyzing isozyme variability among
endosperm, embryo and seed coat samples. Observations
were the following:
PGI: two zones of activity were observed. Variation
was found only in the slowest zone (PGI2) where two
single-banded phenotypes were observed in all but one
C. dupreziana samples while a three-banded phenotype
was observed in all C. sempervirens endosperms used as
control (figure 1). All C. dupreziana endosperms
Table I. Number of studied seeds collected from 6 C.
dupreziana and 1 C. sempervirens trees.
Mother tree code Number of Number of
dissected seeds filled seeds
C. dupreziana D38 200 6 (3%)
D28 184 8 (4.3%)
OO1 173 3 (1.7%)
J1 244 9 (3.7%)
DD5 152 7 (4.6%)
RUS 221 8 (3.6%)
C. sempervirens 12 8 (66%)
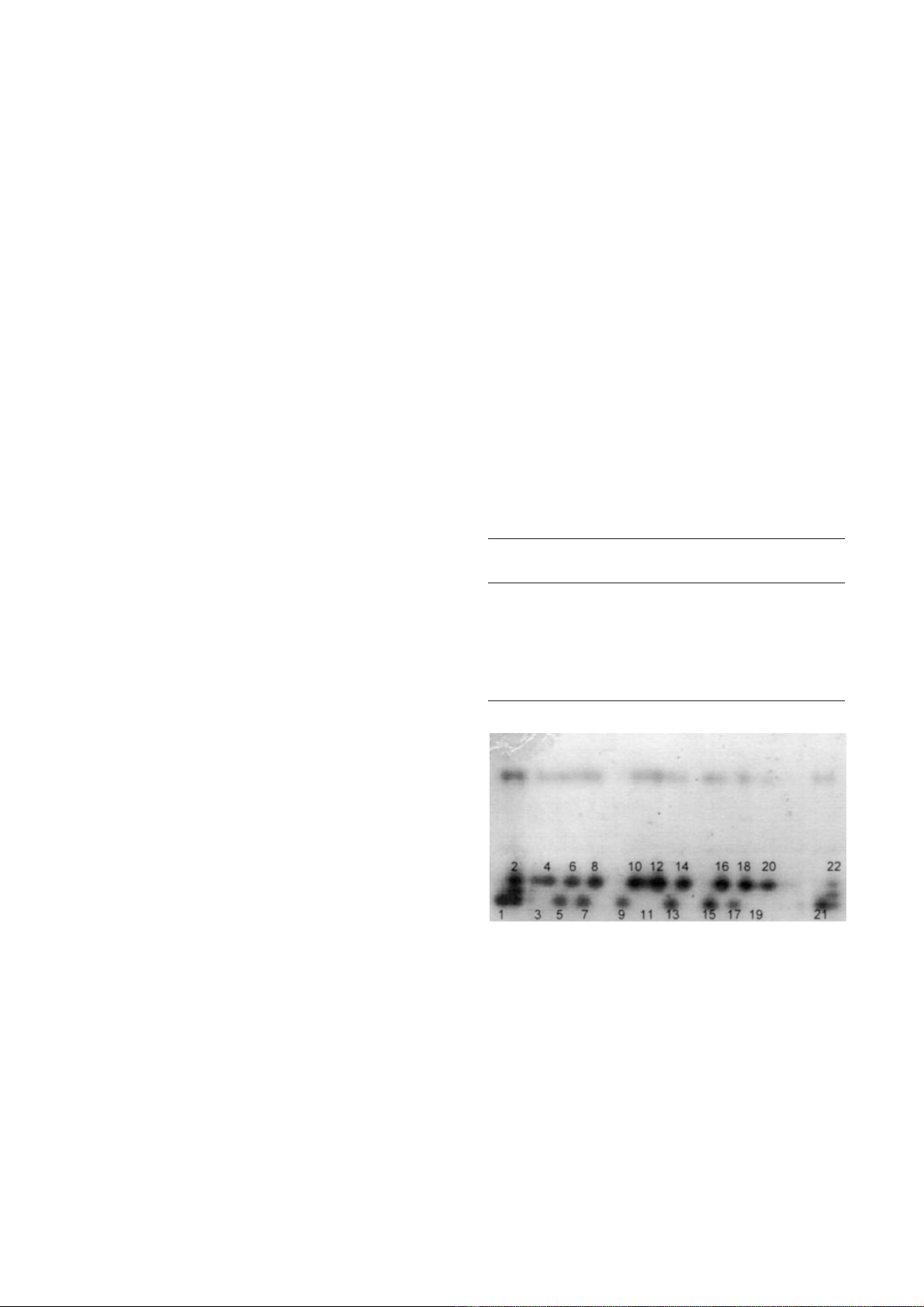
Figure 1. Example of a PGI gel. The monomorphic upper zone
corresponds to PGI1 and the lower polymorphic zone to PGI2.
Samples 1, 2, 21 and 22: C. sempervirens.; all other samples:
C. dupreziana. Embryos: odd samples; corresponding
endosperms: even samples. All C. dupreziana samples have a
single-banded phenotype for PGI2. In 6 of the 9 C. dupreziana
seeds, the endosperm allozyme is absent in the embryo zymo-
gram. C. sempervirens endosperms show a three-banded phe-
notype corresponding to a heterozygous genotype.

Unexpected zymograms in C. dupreziana embryos 19
produced phenotype 1, while embryos produced either
phenotype 1 (8 samples out of 32), phenotype 3 (23/32)
or phenotype 1-2-3 (1/32). For each seed lot, occurrence
of the three observed phenotypes is described in table II.
LAP: one zone of activity was observed. Band 1 was
only observed in C. sempervirens endosperms, while the
3 other bands produced 3 different phenotypes among
the C. dupreziana samples (table III). All C. dupreziana
endosperms produced phenotype 3–4 which was never
observed in embryos. Embryos produced phenotype 2
(22 samples out of 30) or 2–3 (8/30).
4. DISCUSSION
4.1. Empty seeds
The very low percentage of filled seeds observed in
the 6 C. dupreziana seed lots analyzed is in agreement
with previous observations and could be due to abnormal
sexual reproduction [35].
4.2. Genetics of isozyme systems
To our knowledge, there is no published results about
variability of biochemical markers in C. dupreziana,
either with isozymes or with other markers. C. semper-
virens is the closest species for which isozymes studies
are available. Both LAP and PGI enzyme systems were
analyzed in C. sempervirens progenies and/or prove-
nances [18, 21, 28, 30, 31]. Description of zymograms
and inheritance of allozymes were tested by Giannini
et al.[18] using self-pollinated families and by
Papageorgiou et al.[30, 31] using open-pollinated fami-
lies. These authors agreed with the following genetic
interpretation that we considered to be true for C.
dupreziana as well:
PGI2 is a dimeric enzyme with a monolocus
biparental codominant inheritance. Two alleles produced
three phenotypes: 2 single-banded phenotypes for
homozygotes and one three-banded phenotype for het-
erozygotes. In our study, all bands were observed.
LAP is a monomeric enzyme with a monolocus
biparental codominant inheritance. Six bands corre-
sponding to six alleles were revealed. Thus homozygotes
exhibited only one band while heterozygotes exhibited
two bands. In our study, only four bands were observed
and interpreted as 4 alleles.
Table II. Number of samples belonging to one of the 3 PGI2
phenotypes observed. Phenotype numbering refers to decreas-
ing band mobility.
Number of samples per phenotype
Sample 1 3 1-2-3 unreadable
Mother tree Tissue
D38 embryo 0 6 0 0
endosperm 6 0 0 0
D28 embryo 2 5 1 0
endosperm 8 0 0 0
OO1 embryo 1 2 0 0
endosperm 3 0 0 0
J1 embryo 3 6 0 0
endosperm 9 0 0 0
DD5 embryo 2 4 0 0
endosperm 6 0 0 1
C. sempervirens
embryo 0 3 2 0
endosperm 0 0 5 0
Table III. Number of samples belonging to one of the 4 LAP
phenotypes observed. Phenotype numbering refers to decreas-
ing band mobility.
Number of samples per phenotype
Sample 12 2 23 34 unreadable
Mother Tissue
tree
D38 embryo 0 4 2 0 0
endosperm 0 0 0 6 0
D28 embryo 0 4 2 0 2
endosperm 0 0 0 8 0
OO1 embryo 0 1 2 0 0
endosperm 0 0 0 3 0
J1 embryo 0 7 2 0 0
endosperm 0 0 0 9 0
DD5 embryo 0 6 0 0 0
endosperm 0 0 0 6 1
RUS seed coats 0 0 0 4 0
endosperm 0 0 0 8 0
C. sempervirens
embryo 4 4 0 0 0
endosperm 8 0 0 0 0

C. Pichot et al.
20
4.3. Seed tree genotypes in endosperms
Due to the tetrasporic origin of the megagametophyte
[14] and nuclei fusion during its development [13], the
endosperm of C. sempervirens exhibits an enzymatic
phenotype identical to the seed tree phenotype. Thus the
diploid patterns produced by the C.sempervirens
endosperms used as control (tables II and III) show that
this seed tree is heterozygous for both systems LAP and
PGI2.
Endosperms and seed coats extracted from seeds of C.
dupreziana RUS produced an identical two-banded phe-
notypes for LAP, the only system tested. This result sup-
ports the hypothesis of a diploid maternal contribution to
the megagametophyte of C. dupreziana as shown for C.
sempervirens. Thus all C. dupreziana seed trees have the
same genotypes, homozygous for allele 1 in PGI2 and
heterozygous for alleles 3 and 4 in LAP. Such lack of
genotype diversity could be due to the low sample size.
4.4. Embryo genotypes of C. dupreziana seeds
The alleles present in the embryos and absent in the
endosperms (seed trees) indicate the existence of other
genotypic combinations among pollinating trees. Due to
the geographic isolation of the plantation, these alleles
should mainly come from the neighbouring C.
dupreziana trees.
Many embryos analyzed in the present study exhibit-
ed zymograms where band(s) of the corresponding
endosperms were absent: 23 embryos out of 32 for PGI2
and 22 embryos out of 30 for LAP. These two enzymatic
systems were reported to be under nuclear DNA control
with codominant alleles not only for C. sempervirens but
also, to our knowledge, for all gymnosperms studied [1,
7, 8, 12, 15, 19, 22, 25, 32, 37, 38]. All enzymatic sys-
tems tested in C. sempervirens, (including PGI and LAP)
were similarly expressed in endosperm and embryo tis-
sues [29]. The existence of a gene modifying elec-
trophoretic properties, as found for MDH in a few
species [6], has never been demonstrated for PGI and
LAP. Hypothesising, as in C. sempervirens, 1) a nuclear
codominant genetic control, 2) a similar enzymatic tissue
activity and 3) the absence of modifier genes, the fre-
quent lack of maternal alleles in embryo can be best
explain by a strictly paternal origin of embryo nuclear
DNA in C. dupreziana seeds.
In most Cupressaceae studied, such a paternal inheri-
tance was already observed for both chloroplast and
mitichondrial DNA, while in Pinaceae mitochondrial
DNA is usually maternally inherited [9, 26]. But, to our
knowledge, paternal inheritance of the whole nuclear
DNA was never reported.
4.5. Androgenesis and/or paternal apomixis
Paternal inheritance of embryo nuclear DNA implies
embryogenesis from nuclei of the male prothallus that is
produced by germination of a pollen grain.
Production of haploid embryos from anther culture is
a well known phenomenon, called androgenesis [5, 16],
and frequently used in plant breeding. In planta androge-
nesis would imply the production of haploid (or dihap-
loid) embryos. However, a previous study revealed that
C. dupreziana embryos were diploid [35] and some of
the enzymatic phenotypes observed in the present study
corresponded to heterozygous embryos. A “paternal sex-
ual reproduction” could also be hypothesized, where
nuclei fusion occurs between male gametes produced by
one or several pollen grains. However, recent analyses
revealed that C. dupreziana pollen contains unreduced
diploid nuclei [34]. This unknown feature in gym-
nosperms is in agreement with previous morphological
observations on the extraordinary large [17] or aberrant
[2] size of C. dupreziana pollen. It is the largest
observed pollen in the genus Cupressus (38 µvs. 28 µ
for C. sempervirens). We thus hypothesize that unre-
duced male gametes are responsible for the development
of diploid embryo in C. dupreziana.
If the definition of apomixis is not limited to embryo
development from maternal tissue, but means asexual
formation of seed, as commonly accepted [27], then C.
dupreziana may be one apomictic species. Apomixis is
frequently observed in angiosperms and mostly in poly-
ploid species. At least 300 species in 35 families (mainly
Gramineae, Asteraceae, Compositae, Rosaceae and
Rutaceae) are concerned [20, 27]. It has never been
observed in in planta reproduction of gymnosperms [24]
although some references report apomictic type devia-
tions from the sexual cycle of conifers [4, 11, 23].
In comparison with “maternal apomixis”, C.
dupreziana embryo development from paternal tissue (or
at least paternal nuclear DNA) and without fertilization,
would be a unique case of “paternal apomixis”. The bio-
logical significance and adaptive “benefits” of this unex-
pected feature are not clear. It may be the expression of a
trait of survival for a species no more able to produce
female gametes, as inferred from the absence of 1C
nuclei in endosperm.
Further observations using molecular markers con-
trolled by nuclear and cytoplasmic DNA, associated with
cytological observations of female and male tissue
development, before and during embryogenesis, should

Unexpected zymograms in C. dupreziana embryos 21
make it possible to determine the exact contribution of
father and mother trees in C. dupreziana seed produc-
tion, and to know if normal fertilisation sometimes
occurs in this species.
Acknowledgements: We thank Guy Bettachini,
André Giai-Checa and Jean Thévenet for collecting C.
dupreziana seeds, and two anonymous reviewers for
helpful comments on the manuscript.
REFERENCES
[1] Adams W.T., Neale D.B., Doerksen A.H., Smith D.B.,
Inheritance and linkage of isozyme variants from seed and veg-
etative bud tissues in coastal Douglas-fir (Pseudostuga men-
ziesii var. menziesii (Mirb) Franco), Silvae Genet. 93 (1990)
153-157.
[2] Allemand P., Relations phylogéniques dans le genre
Cupressus (Cupressaceae), in: Grasso V., Raddi P. (Eds.). Il
cipresso: Malattie e difesa, AGRIMED, Commission of the
European communities, Firenze, 23-24 November 1979,
pp. 51-67.
[3] Balachowsky A.S., Une relique rarissime du Sahara cen-
tral : Le cyprès de Duprez, La Nature 3237 (1955) 20-24.
[4] Bell P.R., Apomictic features revealed in a conifer, Int.
J. Plant. Sci. 155 (1994) 621-622.
[5] Bonet F.J., Azbaid L., Olmedilla A., Pollen embryogen-
esis: atavism or totipotency?, Protoplasma 202 (1998) 115-121.
[6] Breitenbach-Dorfer M., Geburek T., Gene modifies elec-
trophoretic properties of malate dehydrogenase in Norway
spruce (Picea abies (L.) Karst.), Hereditas 122 (1995) 103-108.
[7] Chagala E.M., Inheritance and linkage of allozymes in
Pinus strobus L., Silvae Genet. 45 (1996) 181-187.
[8] Cheliak W.M., Pitel J.A., Inheritance and linkage of
allozymes in Larix laricina, Silvae Genet. 34 (1985) 142-148.
[9] Chesnoy L., L’origine des organites du cytoplasme
embryonnaire chez les gymnospermes, Bull. Soc. Bot. Fr. 134,
Actual. Bot. 2 (1987) 51-56.
[10] Dobr J., Cupressus dupreziana, Threatened Plants
Newsletter 20 (1988) 8.
[11] Durzan D.J., Jokinen K., Guerra M.P., Santerre A.,
Chalupa V., Havel L., Latent diploid parthenogenesis and
parthenote cleavage in egg-equivalents of Norway spruce, Int.
J. Plant. Sci. 155 (1994) 677-688.
[12] Eckert R.T., Joly R.J., Neale D.B., Genetics of isozyme
variants and linkage relationships among allozyme loci in 35
eastern white pine clones, Can. J. For. Res. 11 (1981) 573-579.
[13] El Maâtaoui M., Pichot C., Nuclei and cell fusion cause
polyploidization in the megagametophyte of common cypress,
Cupressus sempervirens L., Planta 208, 345-351.
[14] El Maâtaoui M., Pichot C., Alzubi H., Grimaud N.,
Cytological basis for a tetraspory in Cupressus sempervirens L.
megagametogenesis and its implications in genetic studies,
Theor. Appl. Genet. 96 (1998) 776-779.
[15] Fady B., Conkle M.T., Segregation and linkage of
allozymes in seed tissues of the hybrid Greek fir Abies borisii
regis Mattfeld, Silvae Genet. 41 (1992) 273-277.
[16] Foroughi-Wehr B., Wenzel G., Andro- and partheno-
genesis, in: Hayward M.D., Bosemark N.O., Romagosa I.
(Eds.). Plant breeding: principles and prospects, Chapman and
Hall Ltd., London, 1993, pp. 261-277.
[17] Gaussen H., Les Gymnospermes actuelles et fossiles,
Trav. Lab. Forest., fascicule X, chapXIII, Toulouse, 1968.
[18] Giannini R., Raddi S., Magnani F., Michelozzi M.,
Palumbo M.G., Final report, in: Teissier du Cros E. (Ed.).
Cypress: A flexible tree for the protection of intensive farm-
land and for the production of high quality wood in marginal
forest sites subject to fire risk in Mediterranean regions,
Commission of the European Communities, DGVI FII.3 Air 3-
CT93-1675, 1997.
[19] King J.N., Dancik B.P., Inheritance and linkage of
isozymes in white spruce (Picea glauca), Can. J. Genet. Cytol.
25 (1983) 430-436.
[20] Koltunow A.M., Apomixis: embryo sacs and embryos
formed without meiosis or fertilization in ovules, (The) Plant
cell 5 (1993)1425-1437.
[21] Korol L., Kara N., Isik K., Schiller G., Genetic differ-
entiation among and within natural and planted Cupressus sem-
pervirens L. Eastern Mediterranean populations. Silvae Genet.
46 (1997) 151-155.
[22] Lewandowski A., Burczyk J., Mejnartowicz L.,
Inheritance and linkage in some allozymes in Taxus baccata
L., Silvae Genet. 41 (1992) 342-347.
[23] Minina E.G., Larionova N.A., Physiological aspects of
possible apomixis in conifers, Soviet-Plant-Physiology 26
(1979) 99-105.
[24] Mogie M., The evolution of asexual reproduction in
plants, Chapman and Hall Ltd., London, 1992.
[25] Morgante M., Vendramin G.G., Giannini R.,
Inheritance and linkage relationships of isozyme variants of
Pinus leucodermis Ant., Silvae Genet. 42 (1993) 231-237.
[26] Neale D.B., Marshall K.A., Harry D.E., Inheritance of
choroplast and mitochondrial DNA in incense-cedar
(Calocedrus decurrens), Can. J. For. Res. 21 (1990) 717-720.
[27] Nijs A.P.M. -den, Dijk G.E. -van, Apomixis, in:
Hayward M.D., Bosemark N.O., Romagosa I. (Eds.). Plant
breeding: principles and prospects, Chapman and Hall Ltd.,
London, 1993, pp. 229-245.
[28] Papageorgiou A., Genetische Untersuchungen zur
Züchtung and Generhaltung bei der Mittelmeerzypresse
(Cupressus sempervirens L.) in Griechenland, Dissertation zur
Erlangung des Doktorgrades des Forstwissenschaftlichen,
Fachbereichs der Georg-August-Universität, Gottingen (1995)
p. 138.
[29] Papageorgiou A., Diploid sporophytic tissue in the seed
of Cupressus sempervirens L., Heredity 81 (1998) 586-590.
[30] Papageorgiou A.C., Bergmann F., Gillet E., Hattemer
H.H., Genetic analysis of isoenzyme variation in
Mediterranean cypress (Cupressus sempervirens L.), Silvae
Genet. 42 (1993) 109-111.


























%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)