
Original
article
Allozyme
variation
in
six
native
oak
species
in
Korea
ZS Kim
SW Lee
JO Hyun
1
Department
of
Forest
Resources,
Korea
University,
Seoul
136-701;
2
Department
of
Forest
Resources,
Seoul
National
University,
Suwon
440-100,
South
Korea
Summary —
Allozyme
variation
at
6
loci
was
studied
in
28
populations
of
the
6
oak
species
native
to
Korea:
Quercus
acutissima
Carruth,
Q
aliena
BI,
Q
dentata
Thunb,
Q
mongolica
Fisch,
Q
serrata
Thunb,
and
Q
variabilis
BI.
The
proportion
of
polymorphic
loci
per
population
(P)
averaged
over
the
6
species
was
74.6%.
The
average
number
of
alleles/locus
(A/L)
was
2.26.
The
average
observed
and
expected
heterozygosities
(H
o,
He)
were
0.302
and
0.298,
respectively.
Only
a
small
amount
(7%)
of
the
observed
genetic
variation
appeared
to
be
interpopulational.
Among
the
6
species,
Q
serrata
and
Q
dentata
were
genetically
less
variable
than
the
others.
Three
loci
could
be used
as
markers
for
distinguishing
Q
acutissima
and
Q
variabilis
from
the other
4
species.
Based
on
genetic
identity,
the
6
oaks
were
also
clustered
into
2
groups.
This
approach
yields
results similar
to
the
current
taxo-
nomic
treatment
by
morphological
characteristics.
allozyme
/
genetic
variation
/
Quercus
species
Résumé —
Variabilité
allozymique
chez
6
espèces
de
chênes
indigènes
de
Corée.
La
variabili-
té
allozymique
a
été
étudiée
dans
28
populations
appartenant
à
6
espèces
de
chênes
indigènes
de
Corée
à
partir
de données
issues
de
6
loci :
Quercus
acutissima
Carruth,
Q
aliena
BI,
Q
dentata
Thunb,
Q
mongolica
Fisch,
Q
serrata
Thunb
et
Q
variabilis
BI.
Le
nombre
moyen
d’allèles
(A/L)
était
de
2,26.
Les
hétérozygoties
observées
et
théoriques
(H
o,
He)
étaient
de
0,302
et
0,298
respective-
ment.
La
variabilité
entre
populations
ne
représentait
que
7%
de
la
variabilité
totale.
Parmi
les
6
es-
pèces,
Q
serrata
et
Q
dentata
étaient
les
moins
variables.
Trois
loci
permettaient
de
distinguer
Q
acutissima
et
Q
variabilis
des
4
autres
espèces.
Le
calcul
des
identités
génétiques
a
permis
de
sé-
parer
les
6
espèces
en
2
groupes.
allozyme
/ variabilité
génétique
/Quercus

INTRODUCTION
Quercus
acutissima
Carruth,
Q
aliena
BI,
Q
dentata
Thunb,
Q
mongolica
Fisch,
Q serrata
Thunb,
and
Q
variabilis
BI
are
the
6
native
deciduous
oaks
which
are
dis-
tributed
throughout
Korea
(Lee
CB,
1987;
Yim,
1991).
They
grow
abundantly
as
dominant
trees
both
in
pure
stands
and
mixed
with
other
species.
This
abundance
is
attributed
mainly
to
vigorous
sprouting
ability
and
viability
on
poor
sites.
In
addi-
tion
to
the
important
role
in
forest
ecosys-
tems,
they
are
economically
valuable
es-
pecially
for
fuel
and
structural
wood
(Lee
CB,
1987;
Yim,
1991).
Due
to
indiscriminate
exploitation
of
the
wood
and
poor
silvicultural
treatments,
most
oak
stands
in
Korea
exhibit
very
poor
growth
and
quality.
In regard
to
their
eco-
nomic
and
ecological
potential,
it
is
likely
that
the
importance
of
oak
species
will
in-
crease
in
the
future.
For
these
reasons,
systematic
genetic
studies
on
oaks
are
now
being
carried
out,
but
they
are
still
at
an
early
stage
(Kim
and
Hyun,
1990;
Lee,
1990;
Park,
1991).
The
objective
of
this
study
was
to
com-
pare
the
genetic
variation
among
the
6
oak
species
and
also
among
populations
in
each
species
by
means
of
isozyme
analy-
sis.
Identification
of
marker
genes
that
can
be
used
to
distinguish
between
the
species
and
clarification
of
the
systematic
relation-
ships
among
them
were
also
points
of
in-
terest.
MATERIALS
AND
METHODS
A
total
of
533
mature
individuals
from
28
popula-
tions
of
the
6
species
were
examined.
From
5
to
30
individuals
per
population
were
assayed.
Lo-
cations
and
the
sample
sizes
are
presented
in
table
I.
For
isozyme
analysis,
young
leaves
forced
out
of
dormant
twigs
were
homogenized
in
a
drop
of
extraction
buffer
(50
ml
of
Tris-HCl
buf-
fer,
pH
7.3,
+
0.06
g
of
ethylenediaminetetra-
acetic
acid
(EDTA)
+
0.05 ml
of
merceptoetha-
nol
+
5
g
polyvinylpyrrolidone
(PVP,
mol
wt
40
000))
and
extracts
were
subjected
to
horizon-
tal
starch-gel
(12.5%)
electrophoresis
using
2
buffer
systems.
System
I was
that
reported
by
Poulik
(1957)
with
slight
modifications;
an
elec-
trode
buffer
of
0.063
M
sodium
hydroxide
titrat-
ed
to
pH
8.20
with
0.299
M
boric
acid
and
a
gel
buffer
of
0.076
M
Tris
titrated
to
pH
8.7
with
0.0068
M
citric
acid.
System
II
consisted
of
an
electrode
buffer
of
0.07
M
Tris
titrated
to
pH
7.0
with
0.021
M
citric
acid
and
a
gel
buffer
obtained
from
a
1:9
aqueous
dilution
of
the
electrode
buf-
fer.
System
I was
used
to
resolve
catalase
(CAT),
leucine
aminopeptidase
(LAP),
menadi-
one
reductase
(MNR)
and
phosphoglucoisomer-
ase
(PGI).
System
II
was
used
to
resolve
aconi-
tase
(ACON).
The
Enzyme
Commission
numbers
of
analyzed
enzymes
and
the
number
of
loci

scored
are
listed
in
table
II.
Enzyme
activity
staining
protocols
were
those
of
Conkle
et
al
(1982)
with
slight
modification.
The
inheritance
pattern
of
observed
enzymes
has
already
been
reported
by
Kim and
Hyun
(1990).
The
genotypes
were
scored
in
the
fol-
lowing
manner:
the
fastest
migrating
locus
was
assigned
A
and
the
next
locus
B
and
so
on;
the
fastest
allozyme
at
a
given
locus
was
designat-
ed
’1’
and
the
slower
forms
were
’2’,
’3’
etc.
Allele
frequencies,
percent
of
polymorphic
loci,
the
mean
observed
and
expected
heterozy-
gosities,
genetic
identities
(Nei,
1978),
and
the
phenogram
drawn
by
the
UPGMA
clustering
technique
(Sneath
and
Sokal,
1973)
were
calcu-
lated
and
produced
using
the
BIOSYS-1
pro-
gram
of
Swofford
and
Selander
(1989).
The
amount
of
interpopulational
genetic
vari-
ation
within
Quercus
species
and
populations
was
determined
by
analyzing
genetic
diversity
measures
(H
T,
HS,
D
ST
and
G
ST
)
(Nei,
1973,
1975).
RESULTS
AND
DISCUSSION
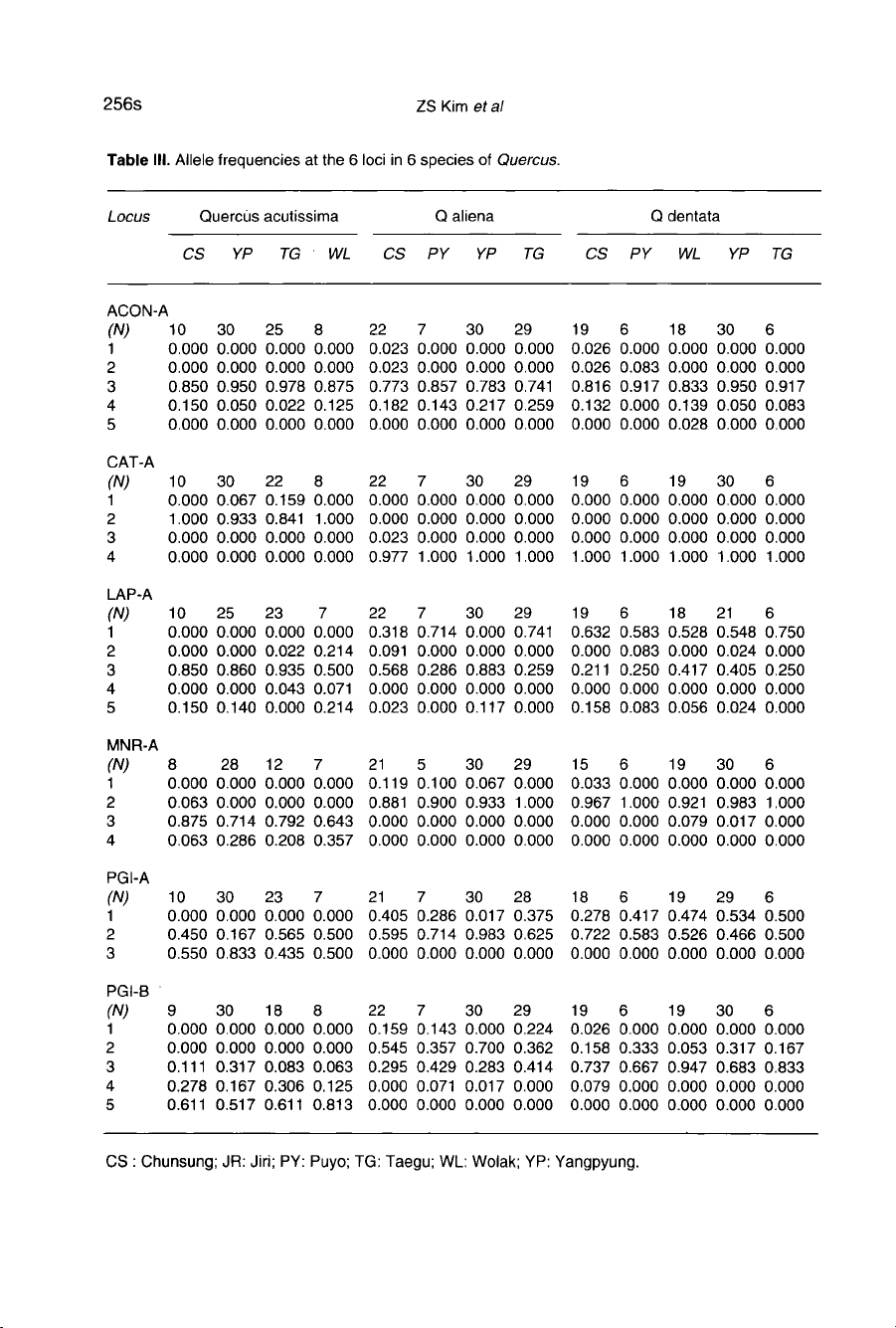
Allele
frequencies
for
the
6
loci
studied
are
presented
in
table
III.
Five
loci
(ACON-A,
LAP-A,
MNR-A,
PGI-A,
PGI-B)
were
poly-
morphic
in
all
6
taxa,
although
not
in
every
population.
Three
loci,
CAT-A,
MNR-A
and
PGI-B,
served
as
allozyme
markers
for
dis-
criminating
Quercus
acutissima
and
Q
vari-
abilis
from
the
other
4
oak
species.
At
CAT-A,
allele
1
was
present
in
Q
acutissi-
ma,
allele
2
in
Q
acutissima
and
Q
variabi-
lis
and
allele
4
was
observed
in
the other
4
species.
Among
the
4
alleles
at
MNR-A,
al-
lele
4
was
found
only
in
Q
acutissima
and
Q
variabilis,
but
allele
1
was
observed
in
the
other
species,
while
alleles
2
and
3
were
displayed
in
all
6
species.
Among
the
5
alleles
at
PGI-B,
allele
5
was
observed
in
Q
acutissima
and
Q
variabilis,
but
alleles
1
and
2
were
found
in
the
other
4
species,
whereas
alleles
3
and
4
were
in
all
6
spe-
cies.
Measurements
of
the
genetic
variability
of
the
6
Quercus
species
are
presented
in
table
IV.
The
percent
of
polymorphic
loci
ranged
from
66.7
(Q
serrata)
to
84.5%
(Q acutissima).
The
amount
of
polymor-
phic
loci
averaged
over
the
6
oak
species
was
74.6%.
The
mean
number
of
alleles/
locus
(A/L)
averaged
over
all
loci
ranged
from
2.03
(Q
serrata)
to
2.65
(Q
mongoli-
ca)
and
the
mean
number
averaged
over
the
6
species
was
2.26.
The
mean
ob-
served
heterozygosities
within
species
ranged
from
0.293
(Q
mongolica)
to
0.307
(Q
aliena
and
Q
serrata).
The
mean
ex-
pected
heterozygosities
ranged
from
0.276
(Q
dentata)
to
0.325
(Q
acutissima).
The
overall
means
of
the
observed
and
expect-
ed
heterozygosities
for
the
6
species
were
0.302
and
0.298,
respectively.


























%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)