
Apoplast:
a
sensitive
site
for
assessing
some
biochemical
effects of
O3
or
SO
2
in
Norway
spruce
needles
G.
Ogier,
F.J.
Castillo
H.
Greppin
Laboratory
of
Plant
Biochemistry
and
Physiology,
University
of
Geneva,
CH-1211,
Geneva
4,
Switzerland
Introduction
The
study
of
the
cell
wall-plasma
mem-
brane
interphase
is
of
great
importance
for
the
understanding
of
gaseous
air
pollu-
tants
and
leaf
cell
interactions.
In
the
apo-
plast
liquid
phase,
the
pollutants
are
solu-
bilized
and
they
can
generate
oxidative
products
(Tingey
and
Taylor,
1982).
For
example,
03
or
S0
2
could
lead
to
H202
production
(Tingey
and
Taylor,
1982;
Khan
and
Malhotra,
1982).
In
order
to
protect
the
plasma
membrane
and
the
compo-
nents
of
the
extracellular
matrix,
cells
are
believed
to
dispose
of
oxidant-scavenging
mechanisms.
One
of
the
enzymatic
sys-
tems
which
could
play
a protective
role
against
oxidative
stresses
includes
per-
oxidases
(Castillo
and
Greppin,
1988).
Peroxidase
activity,
with
guaiacol
as
the
electron
donor,
and
protein
content
were
measured
in
Norway
spruce
needles
(Picea
abies
(L.)
Karst)
after
fumigation
(24
h/d)
in
semi-open
top
chambers,
for
12
wk
in
summer
with
03
or
for
10
wk
in
winter
with
S0
2.
These
parameters
were
followed
in
the
intercellular
washing
fluid
(IWF)
and
in
the
residual
cell
material
(RCM).
The
plants
treated
in
summer
remained
12
wk
longer
in
the
chambers
in
order
to
assess
any
visible
injury
caused
by
03
in
autumn.
Materials
and
Methods
Two
groups
of
20
clone
saplings
(4
yr
old
graft-
ed
P.
abies)
were
selected
from
the
nursery
of
the
Swiss
FedEaral
Institute
of
Forestry
Re-
search
(Birmensdorf,
CH),
one
group
for
each
experiment.
Prior
to
fumigation,
the
plants
were
distributed
randomly
into
4
semi-open
top
chambers
(5
individuals
per
chamber).
Current
year
old
needles
and
1
yr
old
needles
were
analyzed
in
samples
harvested
at
the
end
of
the
fumigation
period.
The
experimental
approach
is
shown
in
Table
I.
The
IWF
was
obtained
after
infiltration
of
phosphate
buffer
(40
mM,
pH
4.5),
0.1
M
KCI,
3
pM
EDTA,
and
centrifugation
(10
000
x
g,
4°C,
10
min)
according
to
Castillo
et al.
(1987).
The
RCM
extract
was
obtained
from
0.5
g
of
the
remaining
needles,
which
were
ground
under
liquid
nitrogen,
in
the
presence
of
PVP
(0.5
g),
then
solubilized
with
3
ml
of
phosphate
buffer
(66
mWl,
pH
7),
and
centrifuged
(10 000 x g, 4°C,
10
min).

Peroxidase
activity
was
assayed
by
mea-
suring
the
oxidation
of
guaiacol
at
470
nm.
This
activity
was
carried
out
using
phosphate
buffer
(66
mM,
pH
6.1),
16
mM
guaiacol,
3.3
mM
H202
and
0-10 ul
of
enzyme
extract.
Protein
contents
were
determined
according
to
Bradford
(1976)
using
a
Bio-Rad
protein
assay
(0-20,ul
of
enzyme
extract).
Statistics.
The
only
environmental
factor
dif-
fering
between
groups
was
the
air
composition
within
the
chambers.
Considering
this
factor,
plants
fumigated
with
either
filtered
air
(fa)
or
fa
plus
added
pollutants
were
under
controlled
conditions
(a);
whereas
in
ambient
air
cham-
bers,
the
fumigation
conditions
were
uncon-
trolled
(b).
In
both
experiments,
the
data
of
the
(a)
plant
groups
were
tested
by
analysis
of
vari-
ance.
Means
which
were
significantly
different
were
identified
using
a
t-test.
The
data
of
the
groups
which
were
statistically
equivalent
were
pooled.
Then,
those
of
the
(b)
plant
groups
were
compared
to
the
pooled
or
unpooled
ones
of
the
(a)
plant
groups
using
a
t-test.
For
these
analyses,
we
chose
P
<0.05
as
significant.
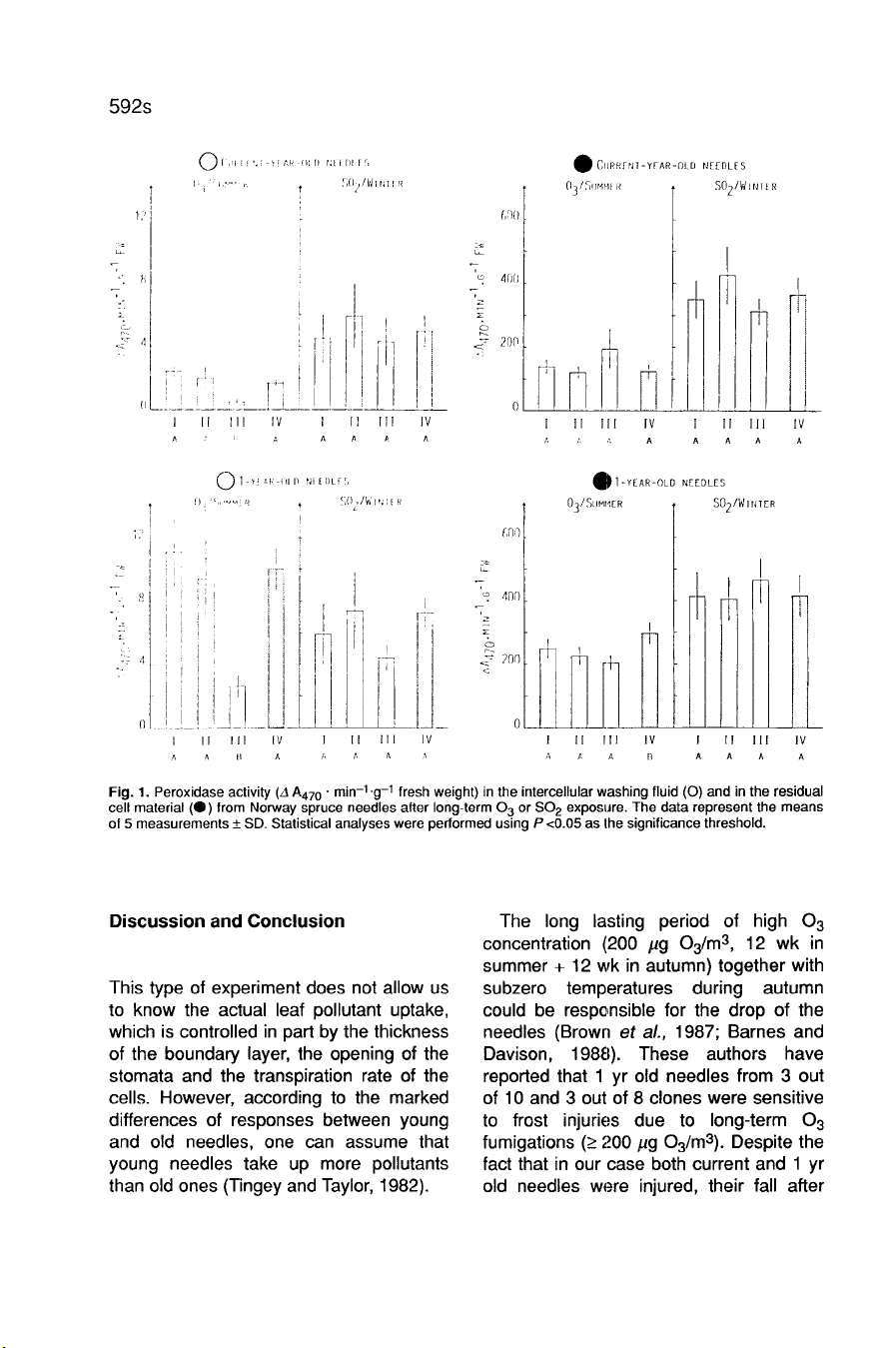
Results
Guaiacol
peroxidase
activity,
(Fig.
1
),
decreased
in
the
IWF
of
current
and
1
yr
old
needles
of
plants
treated
with
200
J1g
Og/m3
(22
and
24%
of
the
control
values,
respectively).
This
enzyme
activity
was
not
affected
in
the
IWF
by
S0
2
treatment.
The
only
noticeable
change
in
the
RCM
was
an
increase
in
1
yr
old
needles
of
the
summer
ambient
air-treated
plants
(124%
of
the
pooled
values
of
the
plants
exposed
to
controlled
conditions).
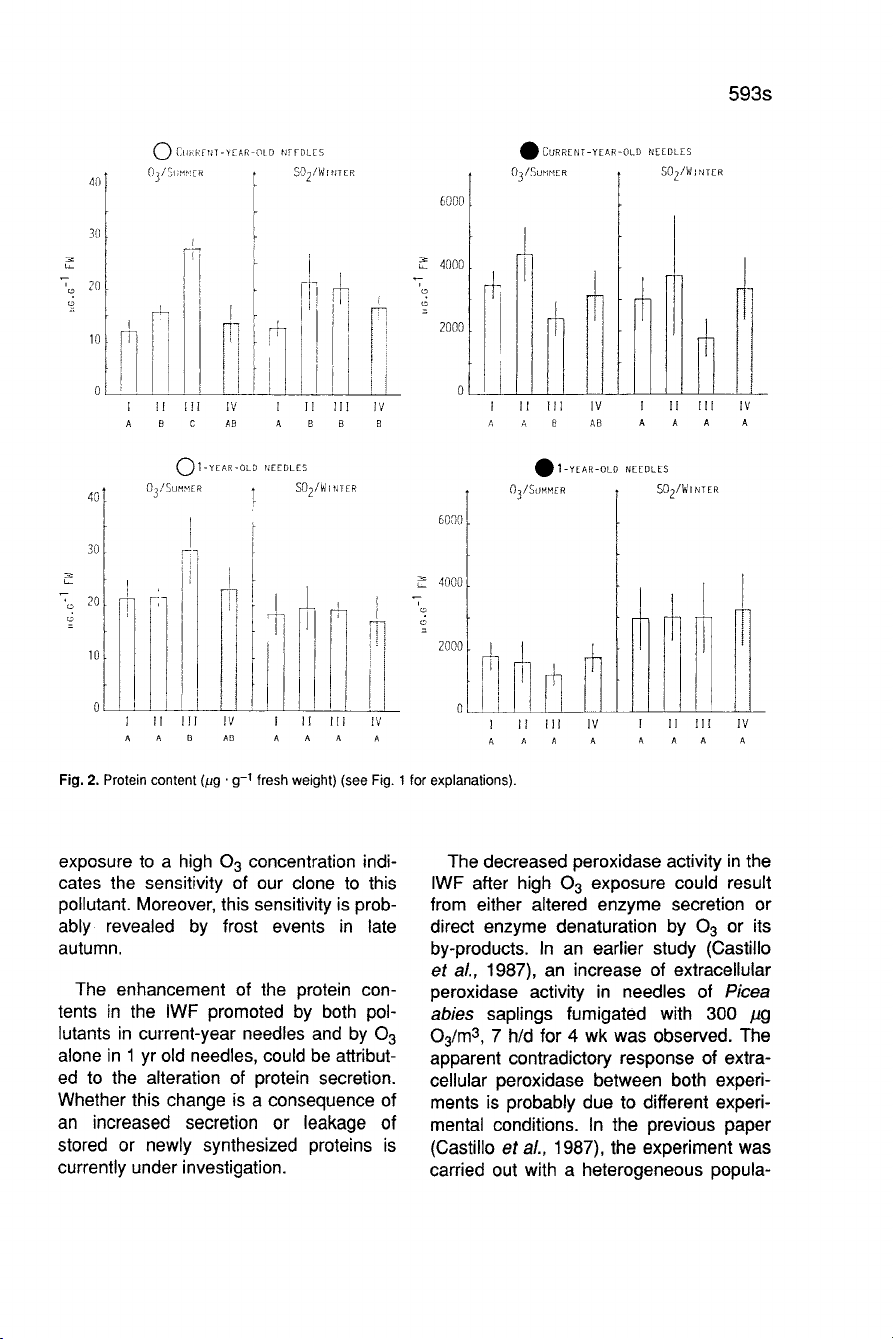
The
protein
content
(Fig.
2)
in
the
IWF
of
young
needles
was
1.3-2.3
times
great-
er
after
low
and
high
ozone
exposure,
and
1.6-1.7
times
greater
after
low
and
high
S0
2
concentrations,
respectively,
as
com-
pared
to
the
control
values.
On
the other
hand,
the
protein
content
of
the
RCM
was
only
affected
by
the
high
ozone
exposure
and
was
lower.
In
the
1
yr
old
needles,
the
only
change
observed
was
an
increase
of
the
protein
content
in
the
IWF
after
high
ozone
exposure.
No
visible
damage
could
be
noted
when
the
plants
were
sampled
either
in
Septem-
ber
(0
3)
or
in
March
(S0
2
).
However,
in
November,
the
needles
of
the
plants
treat-
ed
with
200
!g
03
/m
3
began
to
show
a
’dirty
grey’
aspect
and
to
fall.
By
the
end
of
the
experiment
(late
December)
most
of
the
current
year
and
some
of
the
1
yr
old
needles
were
dead.
This
phenomenon
was
only
observed
in
plants
exposed
to
200 ug
Og/m
3.
Discussion
and
Conclusion
This
type
of
experiment
does
not
allow
us
to
know
the
actual
leaf
pollutant
uptake,
which
is
controlled
in
part
by
the
thickness
of
the
boundary
layer,
the
opening
of
the
stomata
and
the
transpiration
rate
of
the
cells.
However,
according
to
the
marked
differences
of
responses
between
young
and
old
needles,
one
can
assume
that
young
needles
take
up
more
pollutants
than
old
ones
(Tingey
and
Taylor,
1982).
The
long
lasting
period
of
high
03
concentration
(200
!g
03
/m
3,
12
wk
in
summer
+
12
wk
in
autumn)
together
with
subzero
temperatures
during
autumn
could
be
responsible
for
the
drop
of
the
needles
(Brown
et
aL,
1987;
Barnes
and
Davison,
1988).
These
authors
have
reported
that
1
yr
old
needles
from
3
out
of
10
and
3
out
of
8
clones
were
sensitive
to
frost
injuries
due
to
long-term
03
fumigations
(>
200
Pg
03/M3
).
Despite
the
fact
that
in
our
case
both
current
and
1
yr
old
needles
were
injured,
their
fall
after
exposure
to
a
high
03
concentration
indi-
cates
the
sensitivity
of
our
clone
to
this
pollutant.
Moreover,
this
sensitivity
is
prob-
ably
revealed
by
frost
events
in
late
autumn.
The
enhancement
of
the
protein
con-
tents
in
the
IWF
promoted
by
both
pol-
lutants
in
current-year
needles
and
by
03
alone
in
1
yr
old
needles,
could
be
attribut-
ed
to
the
alteration
of
protein
secretion.
Whether
this
change
is
a
consequence
of
an
increased
secretion
or
leakage
of
stored
or
newly
synthesized
proteins
is
currently
under
investigation.
The
decreased
peroxidase
activity
in
the
IWF
after
high
03
exposure
could
result
from
either
altered
enzyme
secretion
or
direct
enzyme
denaturation
by
03
or
its
by-products.
In
an
earlier
study
(Castillo
et
al.,
1987),
an
increase
of
extracellular
peroxidase
activity
in
needles
of
Picea
abies
saplings
fumigated
with
300
Jig
03/M3,
7
h/d
for
4
wk
was
observed.
The
apparent
contradictory
response
of
extra-
cellular
peroxidase
between
both
experi-
ments
is
probably
due
to
different
experi-
mental
conditions.
In
the
previous
paper
(Castillo
et
al.,
1987),
the
experiment
was
carried
out
with
a
heterogeneous
popula-

tion
of
saplings
and
the
total
dose
for
that
short-term
03
fumigation
was
30
ppm/h.
In
this
report,
the
data
were
obtained
from
grafted
saplings
originating
from
the
same
clone
and
the
total
dose
for
this
long-term
03
fumigation
was
200
ppm/h.
Apparently,
extracellular
peroxidase
responds
in
a
dif-
ferent
way
depending
upon
the
level
and
length
of
pollutant
exposure
and/or
on
the
genetic
characteristics
of
the
plant
mate-
rial.
In
the
case
of
high
03
exposure,
the
decreased
extracellular
enzyme
activity
and
the
increased
protein
content
in
the
IWF
of
young
needles
could
be
explained
by
the
high
03
concentration
applied
(200 ,ug
03/M3,
24
hid,
for
12
wk),
which
is
probably
above
the threshold
value
that
the
plant
can
tolerate
without
disruption
of
homeostasis.
Based
on
these
observations,
it
ap-
pears
that
the
apoplast
of
Norway
spruce
needles
is
a
sensitive
site
for
the
detection
of
stresses
induced
by
gaseous
pollutants.
Acknowledgments
This
work
was
supported
by
Grant
Number
4.849.0.85.14
from
the
Swiss
FNRS.
References
Barnes
J.D.
&
Davison
A.W.
(1988)
The
influ-
ence
of
ozone
on
the
winter
hardiness
of
Nor-
way
spruce
(Picea
abies
(L.)
Karst.).
New
Phy-
tol.
108, 159-16E!
Bradford
M.
(1976)
A
rapid
and
sensitive
method
for
the
quantitative
determination
of
microgram
quantities
of
protein
utilizing
the
principle
of
protein-dye
binding.
Anal.
Bio-
chem.
72,
248-2.54
Brown
K.A.,
Rot>erts
TM.
&
Blank
L.W.
(1987)
Interaction
between
ozone
and
cold
sensitivity
in
Norway
spruce:
a
factor
contributing
to
the
forest
decline
in
central
Europe?
New
PhytoL
105, 149-155
Castillo
F.J.
&
Greppin
H.
(1988)
Extracellular
ascorbic
acid
and
enzyme
activities
related
to
ascorbic
acid
metabolism
in
Sedum
album
leaves
after
ozone
exposure.
Environ.
Exp.
Bot. 28, 231-238
Castillo
F.J.,
Miller
P.R.
&
Greppin
H.
(1987)
Extracellular
biochemical
markers
of
photo-
chemical
oxidant
air
pollution
damage
to
Nor-
way
spruce.
Experientia
43, 111-115
s
Khan
A.A.
&
M:alhotra
S.S.
(1982)
Peroxidase
activity
as
an
indicator
of
S0
2
injury
in
jack
pine
and
white
birch.
Biochem.
Physiol.
Pflanz.
177,
643-650
Tingey
D.T.
&
T
ayior
G.E.
(1982)
Variation
in
plant
response
to
ozone:
a
conceptual
model
of
physiological
events.
In:
Effects
of
Gaseous
Air
Pollution
in
Agriculture
and
Horticulture.
Butter-
worths,
London,
pp.
111-138
















%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)