
Minireview
Gene expression neighborhoods
Brian Oliver, Michael Parisi and David Clark
Address: Laboratory of Cellular and Developmental Biology, National Institute of Diabetes and Digestive and Kidney Diseases, National
Institutes of Health, Bethesda, MD 20892, USA.
Correspondence: Brian Oliver. E-mail: oliver@helix.nih.gov
Reductionist approaches have been a tremendous boon to
understanding the regulation of transcription, one of the
vital steps defined by the central dogma of molecular
biology. Gene-by-gene analysis has clearly shown that
control regions within the DNA sequence bind protein
transcription factors that up- or down-regulate the activity
of promoters. But now that patterns of gene expression can
be studied across the entire genome, new findings suggest
that, as well as being controlled individually, genes may
also be subject to regulation according to their location
within the genome.
It has been clear for some time that genomic location has
some impact on gene expression. For example, in various
species when transgenes are removed from their local envi-
ronment and reinserted elsewhere in the genome the trans-
genes tend to work more-or-less normally but almost always
show some alteration in expression due to insertion site - and
sometimes the effect on expression is dramatic. That even
subtle differences in gene expression can have consequences
in some circumstances is also well known, and is illustrated
by the dramatic effects of minute concentration differences
in the gradients of pattern-determining morphogens during
development [1], and in the dosage compensation mecha-
nisms that have evolved to ensure that X-linked genes are
expressed at similar levels in male and female animals [2].
In this issue, Spellman and Rubin [3] describe a transcrip-
tional profiling study that reveals a surprising correlation
between the organization of genes along Drosophila chro-
mosomes and their expression levels. Specifically, neigh-
borhoods composed of an average of 15 contiguous genes
show markedly similar relative expression levels. Although
the average neighborhood contains 15 genes, there is a very
wide range. These neighborhoods are not obviously com-
posed of genes with related functions that might be
expected to exhibit co-regulation, as is the case for the
rRNA, histone, Hox, and globin gene clusters.
Two other recent papers also suggest that genes with similar
expression levels are non-randomly distributed, in this case
within the human genome [4,5]. In humans, it has been sug-
gested recently that expression neighborhoods serve to regu-
late housekeeping functions [5]. In Drosophila this is less
likely, however, because Spellman and Rubin [3] dem-
onstrate that embryos and adults differ dramatically in the
organization of their neighborhoods of similarly expressed
genes (although one could argue about whether the vermi-
form Drosophila larvae and adults might be expected to show
two different housekeeping gene sets). The compelling and
intriguing Drosophila data are rather mysterious and warrant
closer examination: what could underlie the observed
similarity of gene expression within neighborhoods?
Published: 1 July 2002
Journal of Biology 2002, 1:4
The electronic version of this article is the complete one and can be
found online at http://jbiol.com/content/1/1/4
© 2002 BioMed Central Ltd ISSN 1475–4924
Abstract
The finding that neighboring eukaryotic genes are often expressed in similar patterns suggests
the involvement of chromatin domains in the control of genes within a genomic neighborhood.
BioMed Central
Journal
of Biology
Journal of Biology 2002, 1:4
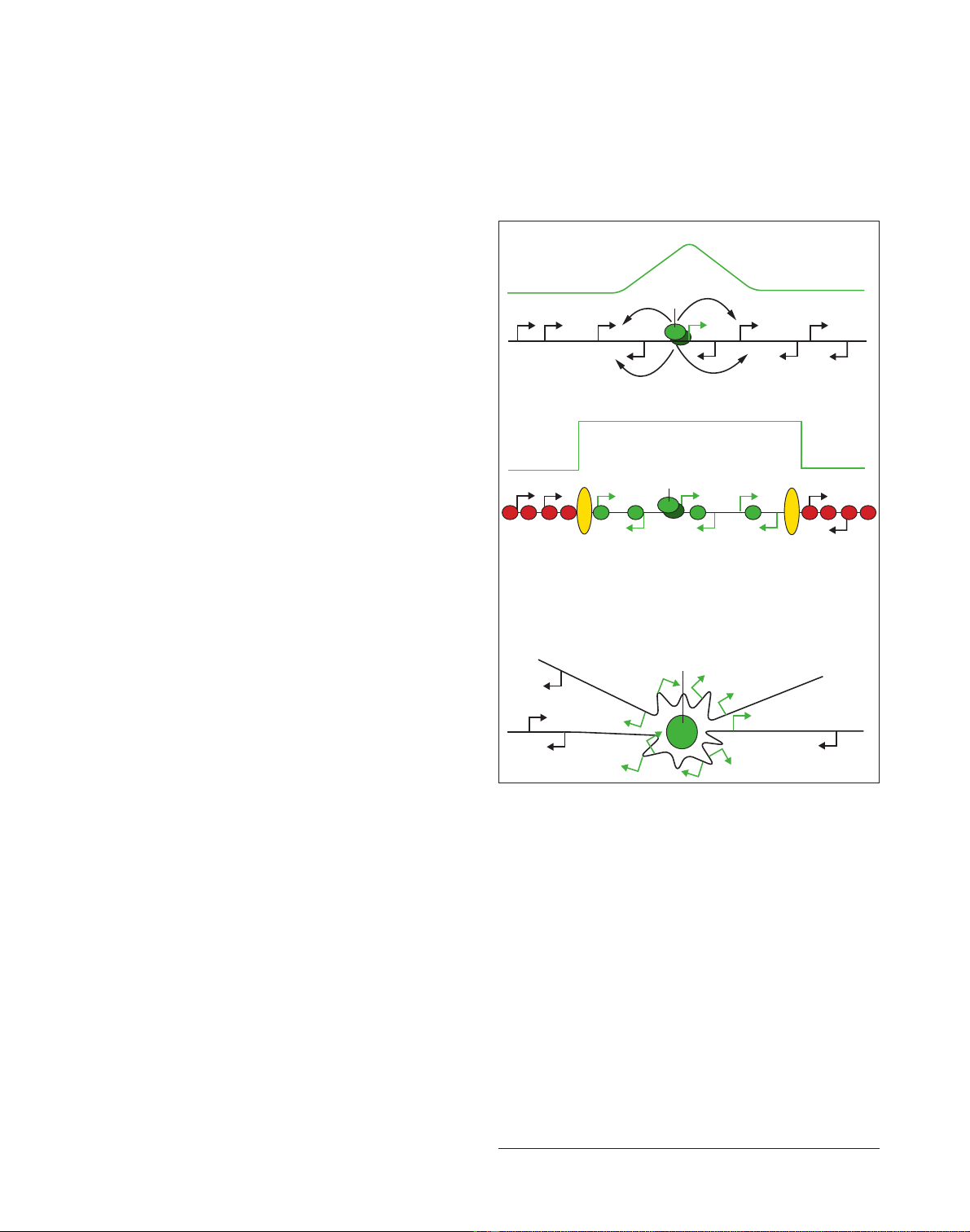
Perhaps the simplest explanation is that co-regulation
within an expression neighborhood may be due to inciden-
tal interactions between promoters and transcriptional
enhancers (Figure 1a). In this model, transcription of one
or more genes in a genomic cluster is regulated by the
usual suspects (transcription factors) binding at the appro-
priate sites and activating nearby genes as well as the
target gene - and the resulting inappropriate expression of
genes other than the target is tolerated because it has little
biological effect. If this is the case then, if sites that bind
strong transcriptional activators, such as the yeast protein
GAL4, were seeded in the Drosophila genome they should
create new neighborhoods. Transcription factors have a
limited range of effect [6], so if strong activators are
responsible one might expect to see a steep fall-off in the
effects of a given factor with distance from its core binding
site (Figure 1a). But the data presented by Spellman and
Rubin [3] suggest that in fact the pattern of gene expres-
sion within a neighborhood is essentially a square wave
(as shown in Figure 1b).
Spellman and Rubin [3] therefore favor a structural chro-
matin domain model (Figure 1b), involving the opening of
the chromatin of an entire neighborhood as a result of acti-
vation of a target gene within the neighborhood. The cre-
ation of a domain of open chromatin structure would, it is
argued [3], increase the availability of the promoters and
enhancers of all the genes in the neighborhood to the tran-
scriptional machinery, leading to correlated increases in
expression. Such a domain could be delimited by boundary
elements or insulators, accounting for the square wave
profile (Figure 1b). A problem with this model is that
increased chromatin accessibility is just as likely to facilitate
the binding of repressors as activators, with the result that
some genes would be up-regulated and some down-regu-
lated. This is not consistent with neighborhoods of co-regu-
lation. But if increased accessibility primarily affects basal
(that is, non-activated) expression, there could be a general
increase in transcription of all the genes in the neighbor-
hood. Indeed, modification of the chromatin of the male X
chromosome in Drosophila results in global up-regulation
of gene expression [2], as does depleting histones from yeast
[7]. And if neighborhoods influence all genes within them -
and not just those that evolved so as to be regulated within a
particular neighborhood - then inserted transgenes that
land in a neighborhood should come under neighborhood
control, and chromosome deletions and inversions should
alter the extent of particular neighborhoods.
Spellman and Rubin [3] tested a short list of known
chromosomal structures to look for correlations with
expression neighborhoods. The cytology of Drosophila
chromosomes and chromosome puffs has long suggested
that the chromosome is divided into loop domains with
differing degrees of compaction. Indeed, heterochromatin
and euchromatin were recognized long before we knew that
4.2 Journal of Biology 2002, Volume 1, Issue 1, Article 4 Oliver et al. http://jbiol.com/content/1/1/4
Journal of Biology 2002, 1:4
Figure 1
Models to account for gene expression neighborhoods. Several models
(or combinations of models) could account for the observed
phenomenon of gene expression neighborhoods. (a) Incidental
regulation. A transcription factor (green oval) binds at a target gene
(green arrow) and incidentally up-regulates neighboring genes. In this
model, the level of expression of neighboring genes is determined by
proximity to the target gene and is expected to decrease with distance
from the target gene (the green line at the top of each panel indicates
the gene expression profile across the neighborhood). (b) A structural
domain model. A discrete ‘open’ chromatin domain is created as a
result of activation of a target gene within the domain. Flanking
boundary or insulator elements (yellow ovals) define the neighborhood
and the limits of the open chromatin domain. (Note the ‘square wave’
expression profile.) (c) Expression neighborhoods in three-dimensional
space. In this model, activation of a target gene results in its
recruitment to a specific nuclear location. This would necessarily
involve the co-recruitment of neighboring genes. The particular
subnuclear location exposes the neighborhood to increased
concentrations of components of the transcriptional machinery (the
image shows two segments of chromatin with two neighborhoods in
the vicinity of a (green) nuclear body).
(a) Incidental expression
(c) Expression neighborhoods in three-dimensional space
Transcription
factor
Transcription
factor
'Closed'
chromatin
'Open' chromatin
'Closed'
chromatin
Boundary Boundary
(b) Structural domain
Nuclear body?

chromosomes were the carriers of genetic information. Mol-
ecular biologists know that chromatin has various accessi-
bility states and binds to a nuclear matrix at defined
locations. Which of these is the structural basis of a neigh-
borhood? The short and surprising answer appears to be
none of the above. Although the stunning block-like orga-
nization of neighborhoods along a chromosome [3] indi-
cates that there must be cis-acting structures, no known
structures correlate with the blocks. But it is increasingly
clear that the nucleus is a highly organized three-dimen-
sional space (Figure 1c). Sub-nuclear structures of various
types, such as insulator bodies and the PML macromolecu-
lar bodies found in mammalian nuclei, may be distinct from
structural elements such as loop-domain boundaries and
matrix-attachment regions [8,9]. The hunt for the structural
basis of expression neighborhoods will be an exciting one.
What do expression neighborhoods mean for the organism?
One possibility, favored by Spellman and Rubin [3], is that
they mean nothing. They suggest that although expression
domains reveal some sort of structural feature, only one or
a few genes in the neighborhood are bona fide targets. The
bottom line for any would-be gene-expression profiler is
that the interesting genes identified in a microarray
experiment are accompanied by a large amount of chaff.
Spellman and Rubin suggest that the inappropriate
expression of gene neighbors does no harm, an idea that is
supported by the lack of dominant phenotypes when
single genes are mutated. But it is also true that deletions
removing greater than 1% of the Drosophila genome
(around 140 genes) have severe dominant deleterious
effects on the organism [10]. Such deletions are likely to
remove whole neighborhoods.
It seems to us that expression neighborhoods should
greatly favor the evolution of genes that benefit by being
within that neighborhood. For example, a de novo function
that is encoded in a gene is of no consequence if it is never
expressed in a tissue that it could influence. As pointed out
by Spellman and Rubin [3], the sequencing of related
Drosophila species will allow us to determine whether
neighborhood structures are maintained intact through
evolutionary time. If the neighborhoods identified by
Spellman and Rubin are less often broken by inversions
than other non-neighborhood regions of the genome
(assuming that there are indeed any non-structured
regions), then neighborhoods are likely to be functionally
significant. Expression neighborhoods could help create,
capture and maintain gene function within a framework of
expression defined by that neighborhood, providing evolu-
tion with additional tools with which to work. From this
fascinating starting point we can expect further insights
into the significance of gene-expression neighborhoods
and the mechanisms that generate them as more genomes
are sequenced and more expression patterns studied over
coming months.
References
1. Tabata T: Genetics of morphogen gradients. Nat Rev Genet
2001, 2:620-630.
2. Pannuti A, Lucchesi JC: Recycling to remodel: evolution of
dosage-compensation complexes. Curr Opin Genet Dev 2000,
10:644-650.
3. Spellman PT, Rubin GM: Evidence for large domains of simi-
larly expressed genes in the Drosophila genome. Journal of
Biology 2002, 1:5.
4. Caron H, van Schaik B, van der Mee M, Baas F, Riggins G, van Sluis
P, Hermus MC, van Asperen R, Boon K, Voute PA, et al.: The
human transcriptome map: clustering of highly expressed
genes in chromosomal domains. Science 2001, 291:1289-
1292.
5. Lercher MJ, Urrutia AO, Hurst LD: Clustering of housekeep-
ing genes provides a unified model of gene order in the
human genome. Nat Genet 2002, 31:180-183.
6. Dorsett D: Distant liaisons: long-range enhancer-promoter
interactions in Drosophila.Curr Opin Genet Dev 1999, 9:505-
514.
7. Wu J, Grunstein M: 25 years after the nucleosome model:
chromatin modifications. Trends Biochem Sci 2000, 25:619-623.
8. Carmo-Fonseca M: The contribution of nuclear compart-
mentalization to gene regulation. Cell 2002, 108:513-521.
9. West AG, Gaszner M, Felsenfeld G: Insulators: many func-
tions, many mechanisms. Genes Dev 2002, 16:271-288.
10. Lindsley DL, Sandler L, Baker BS, Carpenter AT, Denell RE, Hall
JC, Jacobs PA, Miklos GL, Davis BK, Gethmann RC, et al.: Seg-
mental aneuploidy and the genetic gross structure of the
Drosophila genome. Genetics 1972, 71:157-184.
http://jbiol.com/content/1/1/4 Journal of Biology 2002, Volume 1, Issue 1, Article 4 Oliver et al. 4.3
Journal of Biology 2002, 1:4




![Mẫu đề cương chi tiết sinh viên thực tế tại doanh nghiệp [chuẩn nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2014/20140523/duongxuan92/135x160/1681621_356.jpg)











![Báo cáo môn học Công Nghệ Di Truyền [năm] [trường] (hoặc: Mới nhất/Chi tiết)](https://cdn.tailieu.vn/images/document/thumbnail/2013/20130225/trautuongquan/135x160/8601361760920.jpg)


















%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)