
Minireview
The therapeutic potential of modulators of the Hedgehog-Gli
signaling pathway
Barbara Stecca and Ariel Ruiz i Altaba
Address: The Skirball Institute, New York University School of Medicine, 540 First Avenue, New York, NY 10016, USA.
Correspondence: Ariel Ruiz i Altaba. E-mail: ria@saturn.med.nyu.edu
The Hedgehog-Gli signaling pathway regulates numerous
events during the normal development of many cell types
and organs, including the brain, bone, skin, gonads, lung,
prostate, gastrointestinal tract and blood. The hedgehog (hh)
gene - like many of the components of the signaling
pathway triggered by Hedgehog (Hh) protein - was first
identified in Drosophila, where it affects pattern formation
very early in embryonic development. The binding of Hh to
cell membranes triggers a signaling cascade that results in
the regulation of transcription by zinc-finger transcription
factors of the Gli family.
Of the three hh-family genes in mammals - Sonic hedgehog
(Shh), Indian hedgehog (Ihh) and Desert hedgehog (Dhh) -
Shh has been the most studied, mainly because it is
expressed in various tissues but also because experiments
with Shh protein are generally also applicable to other
members of the family. The correct regulation of the
Hh-Gli signaling pathway is essential not only for normal
development but also to prevent a number of human
diseases associated with abnormally increased or
decreased signaling. Here, we discuss the potential use of
small-molecule modulators of the Hh-signaling system,
including those reported by Frank-Kamenetsky et al. in
this issue [1], as therapeutic agents.
Hedgehogs are secreted glycoproteins that act through the
transmembrane proteins Patched1 (Ptc1) and Smoothened
(Smo) to activate an intricate intracellular signal-transduction
pathway (Figure 1). Hh binds Ptc1, a protein with 12 trans-
membrane domains, and this releases the basal repression
that Ptc1 exerts on Smo, a 7-transmembrane-domain protein
that has homology to G-protein-coupled receptors. Inside
the cell, a multimolecular complex, including Costal2
(Cos2), Fused (Fu) and suppressor of Fused (Su(Fu)),
responds to the activation of Smo [2,3] in such a way as to
modify the activity of the Gli proteins (reviewed in [4]).
There are three Gli transcription factors in vertebrates: Gli1
appears to act as a transcriptional activator and is univer-
sally induced in Hh-responding cells, whereas Gli2 and Gli3
can act as activators or repressors of transcription depending
on the particular cellular context. The fate of Gli proteins,
which appear to reside in the cytoplasm in their inactive
state, depends on the state of Hh signaling. In the absence
Published: 6 November 2002
Journal of Biology 2002, 1:9
The electronic version of this article is the complete one and can be
found online at http://jbiol.com/content/1/2/9
© BioMed Central Ltd ISSN 1475–4924
Abstract
The discovery of small molecules that act as agonists and antagonists of the Hedgehog-Gli
signaling pathway, which plays important roles in the embryo and adult, opens a new avenue for
the treatment of diseases caused by aberrant suppression or activation of this complex pathway.
BioMed Central
Journal
of Biology
Journal of Biology 2002, 1:9
of Hh, Gli3 is processed into a smaller, nuclear transcrip-
tional repressor that lacks the carboxy-terminal domain of
full-length Gli3 (Gli-rep in Figure 1). Upon activation of
Smo (and Hh signaling), Gli3 protein cleavage is prevented
and an apparent full-length form with transcription-activat-
ing function is generated (Gli-act in Figure 1). Gli2 also
encodes a repressor function in its carboxy-terminally trun-
cated form, but its formation does not appear to be regu-
lated by Hh signaling.
Mutations in components of the HH-GLI pathway in
humans (human gene and protein names are given in cap-
itals) lead to several diseases that result from either loss of
function or ectopic activation of the pathway (reviewed in
[5]). For example, haploinsufficiency of SHH or mutation
in the human PTCH1 gene are associated with holoprosen-
cephaly, a common syndrome affecting development of
the forebrain and mid-face [6-8]. Moreover, ectopic
expression of Shh, Gli1 or Gli2 in model systems leads to
the formation of tumors that resemble basal cell carcino-
mas (BCCs) ([9-12]; reviewed in [13]), and sporadic
human BCCs consistently express GLI, suggesting that all
sporadic BCCs have this pathway active [10]. Similarly,
human mutations in the Suppressor of Fused - SU(FU) -
gene predispose the carrier to medulloblastoma [14]; spo-
radic medulloblastomas can carry PTCH1 mutations and
express GLI1 - again suggesting that they harbor an active
pathway - and Ptc+/- mice can develop medulloblastomas
([15-19]; reviewed in [13]).
From an examination of the different mutations that cause
aberrant suppression or activation of the HH-GLI pathway
in humans, it seems clear that the development of small
molecules that could act as agonists or antagonists of the
function of proteins such as PTCH1, SMO or GLI might
provide an effective therapeutic approach. One such drug
could be SHH protein itself, a natural agonist. For example, it
has been reported that injection of Shh into the striatum
reduces behavioral deficits in a rat model of Parkinson’s
disease [20], that Shh can induce dopaminergic neuronal dif-
ferentiation [21,22] and that Shh is a neuroprotective agent
[23]. But Shh has a relatively short half-life in serum [24] and
its therapeutic effects have been difficult to evaluate in vivo.
The use of synthetic Hh agonists could therefore provide a
viable alternative to Shh protein. Frank-Kamenetsky et al. [1]
have now identified a synthetic non-peptidyl small mole-
cule that faithfully activates the Hh-Gli pathway, triggering
the known biological effects of Hh signaling. They have
shown that this agonist promotes proliferation and differen-
tiation in a cell-type-specific manner in vitro, while in vivo it
rescues developmental defects of Shh-null mouse embryos.
But this agonist, unlike Shh protein, appears to bypass the
Ptc1-regulatory step, by interacting directly with Smo (see
9.2 Journal of Biology 2002, Volume 1, Issue 2, Article 9 Stecca and Ruiz i Altaba http://jbiol.com/content/1/2/9
Journal of Biology 2002, 1:9
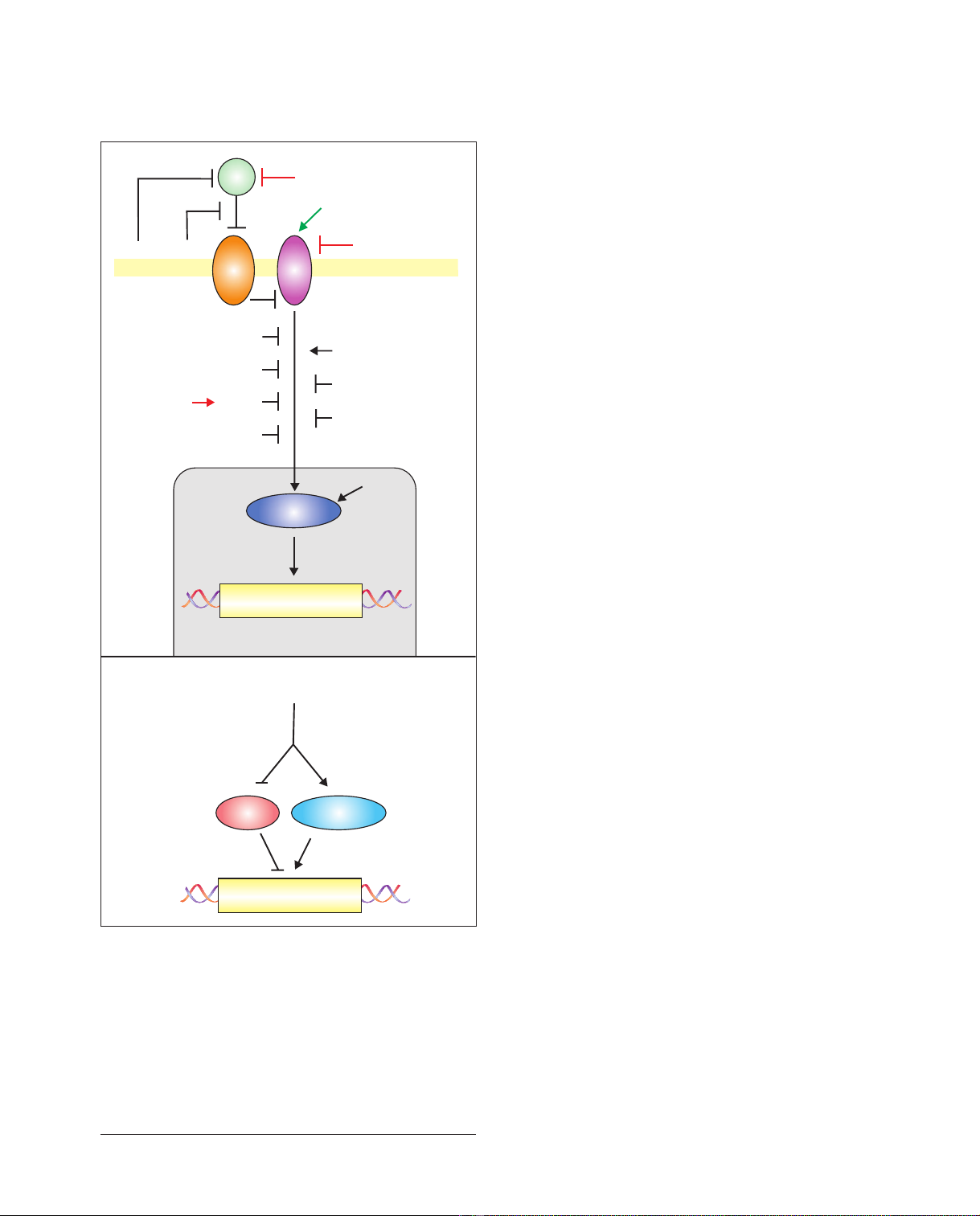
Figure 1
The Hh-signaling pathway. (a) A diagram of the Hh-signaling pathway,
showing the site of action of the agonists (green) and antagonists (red)
discussed in the text, as well as many additional factors that affect the
pathway. Abbreviations: CK1, Casein kinase 1; Cos2, Costal2 ; Dyrk1,
dual-specificity Yak1-related kinase 1; GSK3, Glycogen synthase kinase 3;
Fu, Fused; Gas1, growth arrest specific 1; Hh, Hedgehog; Hip,
Hedgehog-interacting protein 1; Rab23, a Rab-family Ras-like GTPase
associated with vesicle traffic; Ptc, Patched1; PKA, Protein kinase A;
Smo, Smoothened; SuFu, Suppressor of Fused. (b) A schematic
generalized view of the regulation of Gli activator (Gli-act) and Gli
repressor (Gli-rep) forms by Hh signaling. See [2-4] for further details.
Gas1
Ptc Smo
Hip1
Hh
Fu
Hh-Ag
Dyrk1
CK1
GSK3
Anti-Hh antibodies
Cyclopamine
Cur61414
Cos 2
SuFu
PKA
Rab23
Forskolin
Target genes
Nucleus
Membrane
Gli-rep Gli-act
(a)
Gli
Target genes
(b)
Target genes
Hh signaling

Figure 1). Similar results with a near-identical agonist have
now been obtained by another group [25]. From a thera-
peutic point of view, the fact that the molecule retains its
activity after oral administration is a great advantage and,
if its ability to cross the blood-brain and placental barriers
occurs in humans, it could be a very valuable therapeutic
agent. Nevertheless, systemic side effects are to be
expected, as there are many HH-responsive cell populations
in the body.
Treatment of human diseases resulting from ectopic
HH-GLI pathway activation, such as those caused by
oncogenic mutations in SMOH and PTCH1 or in any
element of the pathway that results in activation of GLI
function, requires the use of pathway antagonists. Up to
now, inhibition of ectopic activity has been achieved by
treatment with signaling antagonists that block the
pathway at different levels (Table 1): first, blocking anti-
Shh antibodies that act extracellularly [26]; second,
cyclopamine, a plant alkaloid [27,28] that acts at the level
of Smo in the cell membrane [29]; third, forskolin, an
intracellular activator of protein kinase A (PKA) that is a
cytoplasmic inhibitor of the pathway (see, for example,
[30]); and fourth, Gli-repressor proteins that act within
the nucleus to inhibit positive GLI function from mediat-
ing the HH signal [31] (Figure 1). Therapeutic use of anti-
SHH antibodies is limited to diseases characterized by
misexpression of the ligand and cannot generally be
applied to tumors, because these do not consistently
express SHH (see, for example, [10]). Use of forskolin is
likely to lead to numerous side effects, given the wide-
spread activity of PKA. In contrast, the use of the small
molecule cyclopamine holds great promise.
A number of studies suggest that cyclopamine specifically
inhibits Smo activity [27-29] and that it can affect disease
states caused by activation of the HH-GLI pathway. For
example, the proliferation of a number of human brain-
tumor cell lines and primary tumor cultures, including
those from medulloblastomas and some gliomas [18] as
well as medulloblastoma allografts [32], are inhibited by
treatment with cyclopamine. This suggests that pathway
activation is required for tumor maintenance. Other experi-
ments suggest that the activity of Gli proteins, the terminal
elements of the pathway, is sufficient to induce tumor
development ([10-12]; reviewed in [13]). Thus, HH-
pathway activity may be involved in the initiation as well
as the maintenance of different tumors. This provides an
additional opportunity to inhibit the growth of a number
of tumors in different organs and tissues, such as basal cell
carcinoma in the skin and medulloblastoma in the brain,
with the same agent. Cyclopamine could be such an agent
if the diseases to be treated arise from activation of the HH-
signaling pathway at the level of SMOH or above. In addi-
tion, Frank-Kamenetsky et al. [1] report the use of a new,
synthetic, small-molecule inhibitor, Cur61414, which has
inhibitory properties similar to those of cyclopamine and
also acts at the level of Smo [33]. Whether Cur61414, or
four additional small-molecule antagonists (SANT1-4) that
also act on Smo and were recently identified [25], will
prove to be better and easier to use than cyclopamine
remains to be determined, but testing them against skin
[33] and brain tumors is warranted from a biological point
of view.
Finally, given that carboxy-terminally truncated repressor
forms of GLI3 are potent inhibitors of the activating output
of the HH-signaling pathway [31,34,35], these could be
used as antagonists for the treatment of tumors. The diffi-
culty of delivering them into cells might require the devel-
opment of in vivo transducing strategies, taking advantage,
for example, of the ability of the Penetratin peptide to cross
cell membranes while loaded with cargo [36]. It also sug-
gests that it would be useful to search for and design small
molecules that inhibit GLI’s transcription-activating func-
tion, perhaps by promoting endogenous GLI-repressor for-
mation. This may be very difficult, but such drugs would be
very specific and would be usable in cases where the cancer
is due to mutation in the pathway at any level, from the
extracellular ligand, the HH proteins, to the final mediators,
the GLI proteins.
Agents that inhibit HH signaling may induce the regression
of tumors that are dependent on a deregulated HH-GLI
pathway, but these agents are likely also to affect the
behavior of other normal pathway-dependent cells in the
patient. This may, however, be a small price to pay in
http://jbiol.com/content/1/2/9 Journal of Biology 2002, Volume 1, Issue 2, Article 9 Stecca and Ruiz i Altaba 9.3
Journal of Biology 2002, 1:9
Table 1
Examples of diseases caused by loss of or ectopic function of
the HH-GLI signaling pathway, and the possible agents that
could, in principle, be used as therapeutics
Disease type Potential therapeutic
Gain-of-function: Basal cell carcinoma Antagonist: Anti-HH antibodies
Medulloblastoma Forskolin
Rhabdomyosarcoma Cyclopamine
Cur61414
GLI repressors
Loss-of-function: Holoprosencephaly Agonist: SHH
Hh-Ag*
*Hh-Ag is the Hh agonist described by Frank-Kamenetsky et al. [1].

order to combat cancer, and the agents may have fewer
side effects than current non-specific cytotoxic anti-cancer
chemotherapies.
References
1. Frank-Kamenetsky M, Zhang XM, Bottega S, Guicherit O, Wichterle
H, Dudek H, Bumcrot D, Wang FY, Jones S, Shulok J, Rubin LL,
Porter JA: Small molecule modulators of hedgehog signaling:
identification and characterization of smoothened agonists
and antagonists. J Biol 2002, 1:10.
2. Ho KS, Scott MP: Sonic hedgehog in the nervous system:
functions, modifications and mechanisms. Curr Opin Neuro-
biol 2002, 12:57-63.
3. Nybakken K, Perrimon N: Hedgehog signal transduction:
recent findings. Curr Opin Genet Dev 2002, 12:503-511.
4. Ruiz i Altaba A, Palma V, Dahmane N: Hedgehog-Gli signalling
and the growth of the brain. Nat Rev Neurosci 2002, 3:24-33.
5. Mullor J, Sanchez P, Ruiz i Altaba A: Pathways and conse-
quences: Hedgehog signaling in human diseases. Trends Cell
Biol, in press.
6. Roessler E, Belloni E, Gaudenz K, Jay P, Berta P, Scherer SW, Tsui
LC, Muenke M: Mutations in the human sonic hedgehog
gene cause holoprosencephaly. Nat Genet 1996, 14:357-360.
7. Ming JE, Kaupas ME, Roessler E, Brunner HG, Golabi M, Tekin M,
Stratton RF, Sujansky E, Bale SJ, Muenke M: Mutations in
PATCHED-1, the receptor for SONIC HEDGEHOG, are
associated with holoprosencephaly. Hum Genet 2002,
110:297-301.
8. Muenke M, Beachy PA: Holoprosencephaly. In Metabolic and
Molecular Bases of Inherited Disease. Edited by Scriver CR, Beaudet
AL, Sly WS, Valle D, Childs B, Hilds B, Kinzler KW, Vogelstein B.
New York: McGraw-Hill; 2001:6203-6230.
9. Oro AE, Higgins KM, Hu Z, Bonifas JM, Epstein EH, Scott MP:
Basal cell carcinomas in mice overexpressing sonic hedge-
hog. Science 1997, 276:817-821.
10. Dahmane N, Lee J, Robin P, Heller P, Ruiz i Altaba: Activation of
the transcription factor Gli1 and the Sonic hedgehog sig-
nalling pathway in skin tumours. Nature 1997, 389:876-881.
11. Nilsson M, Unden AB, Krause D, Malmqwist U, Raza K, Zaphi-
ropoulos PG, Toftgard R: Induction of basal cell carcinomas
and trichoepitheliomas in mice overexpressing GLI-1. Proc
Natl Acad Sci USA 2000, 97:3438-3443.
12. Grachtchouk M, Mo R, Yu S, Zhang X, Sasaki H, Hui C-c, Dlugosz
AA: Basal cell carcinomas in mice overexpressing Gli2 in
skin. Nat Genet 2000, 24:216-217.
13. Ruiz i Altaba A., Sanchez P, Dahmane N: Gli and hedgehog in
cancer: tumours, embryos and stem cells. Nat Rev Cancer
2002, 2:361-372.
14. Taylor MD, Liu L, Raffel C, Hui C-c, Mainproze TG, Zhang X,
Agatep R, Chiappa S, Gao L, Lowrance A, et al.: Mutations in
SUFU predispose to medulloblastoma. Nat Genet 2002,
31:306-310.
15. Goodrich LV, Milenkovic L, Higgins KM, Scott MP. Altered
neural cell fates and medulloblastoma in mouse patched
mutants. Science 1997, 277:1109-1113.
16. Raffel C, Jenkins RB, Frederick l, Hebrink D, Alderete B, Fults DW,
James CD: Sporadic medulloblastomas contains PTCH
mutations. Cancer Res 1997, 57:842-845.
17. Wolter M, Reifenberger J, Sommer C, Ruzicka T, Reifenberger G:
Mutations in the human homologue of the Drosophila
segment polarity gene patched (PTCH) in sporadic basal
cell carcinomas of the skin and primitive neuroectoder-
mal tumors of the central nervous system. Cancer Res 1997,
57:2581-2585.
18. Dahmane N, Sanchez P, Gitton Y, Palma V, Sun T, Beyna M, Weiner
H, Ruiz i Altaba A: The Sonic Hedgehog-Gli pathway regu-
lates dorsal brain growth and tumorigenesis. Development
2001, 128:5201-5212.
19. Pomeroy SL, Tamayo P, Gaasenbeek M, Sturla LM, Angelo M,
McLaughlin ME, Kim JY, Goumnerova LC, Black PM, Lau C, et al.:
Prediction of central nervous system embryonal tumour
outcome based on gene expression. Nature 2002, 415:436-
442.
20. Tsuboi K, Shults CW: Intrastriatal injection of sonic hedge-
hog reduces behavioral impairment in a rat model of
Parkinson’s disease. Exp Neurol 2002, 173:95-104.
21. Wang MZ, Jin P, Bumcrot DA, Marigo V, McMahon AP, Wang E,A
Woolf T, Pang K: Induction of dopaminergic neuron pheno-
type in the midbrain by Sonic hedgehog protein. Nat Med
1995, 1:1184-1188.
22. Hynes M, Porter JA, Chiang C, Chang D, Tessier-Lavigne M,
Beachy PA: Induction of midbrain dopaminergic neurons
by Sonic hedgehog. Neuron 1995, 15:35-44.
23. Miao N, Wang M, Ott JA, D’Alessandro JS, Woolf TM, Bumcrot
DA, Mahanthappa NK, Pang K: Sonic hedgehog promotes the
survival of specific CNS neuron populations and protects
these cells from toxic insult in vitro.J Neurosci 1997,
17:5891-5899.
24. Pepinsky RB, Shapiro RI, Wang S, Chakraborty A, Gill A, Lepage
D, Wen D, Rayhorn, Horan GSB, Taylor FR, et al.: Long-acting
form of sonic hedgehog with improved pharmacokinetic
and pharmacodynamic properties are efficacious in a
nerve injury model. J Pharm Sci 2002, 91:371-387.
25. Chen JK, Taipale J, Young KE, Maiti T, Beachy PA: Small mole-
cule modulation of Smoothened activity. Proc Natl Acad Sci
USA 2002, 99:14071-14076.
26. Ericson J, Morton S, Kawakami A, Roelink H, Jessel TM: Two
critical periods of sonic hedgehog signaling required for
the specification of motor neuron identity. Cell 1996,
87:661-673.
27. Incardona JP, Gaffield W, Kapur RP, Roelink H: The teratogenic
Veratrum alkaloid cyclopamine inhibits sonic hedgehog
signal transduction. Development 1998, 125:3553-3562.
28. Cooper MK, Porter JA, Young KE, Beachy PA: Teratogen-
mediated inhibition of target tissue response to Shh sig-
naling. Science 1998, 280:1603-1607.
29. Taipale J, Chen JK, Cooper MK, Wang B, Mann RK, Milenkovic L,
Scott MP, Beachy PA: Effects of oncogenic mutations in
Smoothened and Patched can be reversed by
cyclopamine. Nature 2000, 406:1005-1009.
30. Fan CM, Porter JA, Chiang C, Chang DT, Beachy PA, Tessier-
Lavigne M: Long-range sclerotome induction by sonic
hedgehog: direct role of the amino-terminal cleavage
product and modulation by the cyclic AMP signaling
pathway. Cell 1995, 81:457-465.
31. Ruiz i Altaba: Gli proteins encode context-dependent posi-
tive and negative functions: implication for development
and diseases. Development 1999, 126:3205-3216.
32. Berman DM, Karhadkar SS, Hallahan AR, Pritchard JI, Eberhart
CG, Watkins DN, Chen JK, Cooper MK, Taiplae J, Olson JM,
Beachy PA: Medulloblastoma growth inhibition by hedge-
hog pathway blockade. Science 2002, 297:1559-1561.
33. Williams JA, Guicherit OM, Zaharian BI, Xu Y, Chai L, Gatchalian
C, Porter JA, Rubin LL, Wang FY: Identification of novel
inhibitors of the hedgehog signaling pathway: effects on
basal cell carcinoma-like lesions. Proc Natl Acad Sci USA 2002,
in press.
34. Sasaki H, Nishizaki Y, Hui C, Nakafuku M, Kondoh H: Regula-
tion of Gli2 and Gli3 activities by an amino-terminal
repressor domain: implication of Gli2 and Gli3 as primary
mediators of Shh signaling. Development 1999, 126:3915-
3924.
35. Shin SH, Kogerman P, Lindstrom E, Toftgard R, Biesecker LG:
Gli3 mutations in human disorders mimic Drosophila
cubitus interruptus protein functions and localizations.
Proc Natl Acad Sci USA 1999, 96:2880-2884.
36. Derossi D, Chassing G, Prochiantz A: Trojan peptides: the
penetratin system for intracellular delivery. Trends Cell Biol
1998, 8:84-87.
9.4 Journal of Biology 2002, Volume 1, Issue 2, Article 9 Stecca and Ruiz i Altaba http://jbiol.com/content/1/2/9
Journal of Biology 2002, 1:9
















%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)