92
TỔNG QUAN VỀ ĐÍCH TÁC ĐỘNG TIỀM NĂNG TRÊN CHỦNG
STAPHYLOCOCCUS AUREUS KHÁNG METHICILLIN (MRSA)
Mai Thị Ngọc Lan Thanh1
1. Trường Đại Học Thủ Dầu Một
TÓM TẮT
Ngày nay, vi khuẩn kháng kháng sinh ngày càng phát trin mạnh mẽ, đặc biệt là nhóm vi
khuẩn gồm các chủng Enterococcusfaecium, Staphylococcus aureus, Klebsiella pneumoniae,
Acinetobacter baumannii, Pseudomonas aeruginosa, và Enterobacter spp., trong đó,
Staphylococcus aureus kháng methicllin (MRSA) được xếp vào nhóm quan trọng ưu tiên toàn
cầu đ nghiên cứu và phát trin các phương pháp điều trị mới. MRSA có nhiều cơ chế kháng
thuốc và các đích tác động tiềm năng, bài báo này nhằm tổng quan các đích tác động tiềm năng
làm cơ sở cho việc sàng lọc các hợp chất hay thuốc kháng MRSA gồm màng sinh học, hệ thống
tín hiệu Quorum sensing, độc tố, vách tế bào, cung như kênh bơm thuốc.
Từ khóa: MRSA, đích tác động màng sinh học, độc tính, vách tế bào
1. ĐẶT VẤN ĐỀ
Chi Staphylococcus có hơn 40 loài khác nhau, trong đó có 3 loài tụ cầu gây bệnh được
chú ý trong y học: S. aureus (Tụ cầu vàng), S. epidermidis (Tụ cầu da), và S. saprophyticus. Ở
cơ thể người, S. aureus là vi sinh vật cơ hội tấn công vào vết thương hở hoặc những người có
sức đề kháng kém. Vi khuẩn thường kí sinh trên mũi, họng và da của con người và động vật.
Trong chiến tranh thế giới thứ hai, penicillin được đưa vào sử dụng lâm sàng, S. aureus lúc này
còn rất nhạy với kháng sinh. Tuy nhiên, chỉ vài năm sau đó các chủng S. aureus kháng penicilin
đã được phát hiện (Dien Bard, Hindler, Gold, & Limbago, 2014). Hiện nay, S. aureus kháng
penicillin - PRSA (Penicillin Resistant S. aureus) chiếm khoảng 90 % , MRSA dao động từ 30-
50 % trong tổng số các chủng S. aureus được phân lập (Bộ Y Tế, 2015). Methicilin là penicilin
bán tổng hợp, MRSA kháng methicillin xuất hiện 2 cơ chế kháng phổ biến: một là siêu biểu
hiện β-lactamases, hai là thay đổi dạng bình thường của Protein gắn Penicillin (PBPs). Biểu
hiện enzyme β-lactamase được kiểm soát bởi operon bla. Trong Operon bla, blaZ là vùng gen
mã hóa cho β-lactamase, và hai protein đóng vai trò là các protein điều hòa là: blaI (nhân tố ức
chế) và blaR1 (nhân tố hoạt hóa). BlaI ức chế sự biểu hiện của blaZ bằng việc gắn với promoter.
BlaR1 tồn tại như là protein xuyên màng; khi vùng bên ngoài tế bào gắn với β-lactam, thì vùng
bên trong tế bào sẽ được giải phóng và phân giải BlaI, việc này cho phép biểu hiện blaZ. Cơ
chế kháng thứ hai là biểu hiện protein PBP2a được mã hóa từ gen mecA, nằm trên vùng Operon
mec. Operon mecA có hai nhân tố điều hòa là các protein mecI (là nhân tố ức chế) và mecR1 (
là nhân tố hoạt hóa nằm xuyên màng). mecI và mecR1 có chức năng tương đồng với blaI và
blaR1; việc gắn với β-lactam, vùng mecR1 bên trong tế bào được giải phóng để phân giải mecI
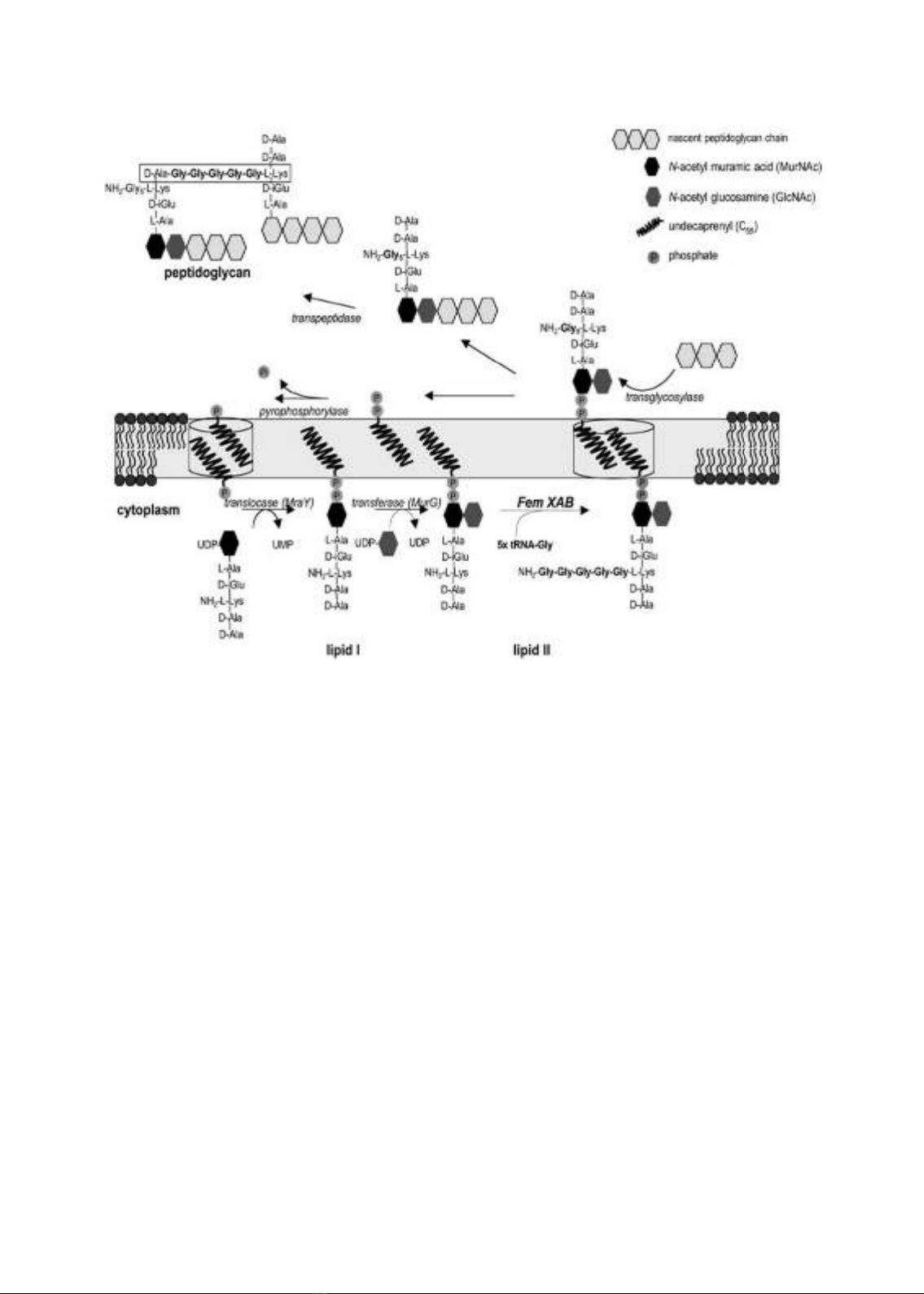
và cho phép sự phiên mã mecA. Sự hiện diện của protein PBP2a-là cơ chế hiện diện sau cùng
trong hầu hết các chủng lâm sàng. S. aureus có 4 tiền chất PBPs thông thường trên màng tế bào
chất tham gia vào quá trình liên kết chéo peptidoglycan của vách tế bào. Những PBPs này có
hoạt động tương tự với serine của protease và có ái lực cao với những chất β-lactam. Khi xảy
ra việc gắn, những PBPs không có chức năng hình thành phức hợp vách tế bào, dẫn đến vi