
A thermoacidophilic endoglucanase (CelB) from
Alicyclobacillus
acidocaldarius
displays high sequence similarity to
arabinofuranosidases belonging to family 51 of glycoside
hydrolases
Kelvin Eckert and Erwin Schneider
Humboldt Universita
¨t zu Berlin, Institut fu
¨r Biologie/Bakterienphysiologie, Berlin, Germany
A 100-kDa protein with endoglucanase activity was purified
from Triton X-100 extract of cells of the thermoacidophilic
Gram-positive bacterium Alicyclobacillus acidocaldarius.
The enzyme exhibited activity towards carboxy methyl cel-
lulose and oat spelt xylan with pH and temperature optima
of 4 and 80 C, respectively. Cloning and nucleotide
sequence analysis of the corresponding gene (celB) revealed
an ORF encoding a preprotein of 959 amino acids which
is consistent with an extracellular localization. Purified
recombinant CelB and a variant lacking the C-terminal 203
amino acid residues (CelB
trunc
) displayed similar enzymatic
properties as the wild-type protein. Analysis of product
formation suggested an endo mode of action. Remarkable
stability was observed at pH values between 1 and 7 and
60% of activity were retained after incubation for 1 h at
80 C. CelB displayed homology to members of glycoside
hydrolase family 51, being only the second entry with activity
typical of an endoglucanase but lacking activity on p-nitro-
phenyl-a-
L
-arabinofuranoside (pNPAraf). Highest sequence
similarity was found towards the other endoglucanase F
from Fibrobacter succinogenes (EGF), forming a distinct
group in the phylogenetic tree of this family. Analysis of the
amino acid composition of the catalytic domains demon-
strated that CelB contains fewer charged amino acids than
its neutrophilic counterparts, which is in line with adaptation
to low pH. Wild-type and full-length recombinant CelB were
soluble only in Triton X-100. In contrast, CelB
trunc
was
completely water soluble, suggesting a role of the C-terminal
region in cell association. This C-terminal hydrophobic
region displayed local sequence similarities to an a-amylase
fromthesameorganism.
Keywords: endoglucanase; EC 3.2.1.4; enzyme 1,4-b-
D
-glu-
can glucanohydrolase; glycoside hydrolase family 51;
acidophile; Alicyclobacillus.
Cellulose and hemicellulose (e.g. xylan), the major compo-
nents of the plant cell wall, constitute complex substrates as
variation can occur in the nature of the monomers, the
linkages, chain length and degree of substitution. The
complexity and variety of these substrates are mirrored by
the numerous enzymes employed by microorganisms to
degrade them. Thus, conversion of cellulose and xylan to
soluble products requires endoglucanases (1,4-b-
D
-glucan-
4-glucanohydrolase; EC 3.2.1.4), exoglucanases, including
cellodextrinases (1,4-b-
D
-glucan glucanohydrolase; EC
3.2.1.74) and cellobiohydrolases (1,4-b-
D
-glucan cello-
biohydrolase; EC 3.2.1.91), b-glucosidases (b-glucoside
glucohydrolase; EC 3.2.1.21), xylanases (1,4-b-
D
-xylan
xylanohydrolase; EC 3.2.1.8) and b-xylosidases (1,4-b-
D
-
xylan xylohydrolase; EC 3.2.1.37) [1]. To reflect structural
features and to reveal the evolutionary relationships
between these enzymes, glycoside hydrolases have been
grouped into families on the basis of sequence similarity [2].
Some families contain enzymes with different substrate
specificities while, on the other hand, enzymes with the same
activity are found in different families [3]. Thus, cellulases
are found in families 5–10, 12, 44, 45, 48, 51, 61 and 74, while
xylanases have been assigned to families 10, 11, and 43.
Cellulolytic and xylanolytic activities are also widespread
in thermophilic microorganisms. Their occurrence is testi-
mony to the presence of these substrates in thermophilic
environments, either as plant litter in natural hot springs or
in environments such as composite piles. Remarkably
however, with a few exceptions, degradation of cellulose
and hemicellulose among thermophiles is mostly due to
anaerobic species and is absent in archaea [4]. Endoglucan-
ases from aerobic thermophilic bacteria, displaying tem-
perature optima between 55 and 70 C and pH optima
between 5 and 7 have been described so far for Acidother-
mus cellulolyticus [5], Rhodothermus marinus [6], Thermobi-
fida fusca [7], and Caldibacillus cellulovorans [4]. Based on
16
S-rRNA gene sequence, the latter is a close relative of
members of the genus Alicyclobacillus that is characterized
by the presence of alicyclic fatty acids as major components
Correspondence to E. Schneider, Humboldt Universita
¨t zu Berlin,
Institut fu
¨r Biologie/Bakterienphysiologie, Chausseestr. 117,
D-10115 Berlin, Germany.
Fax: + 49 (0)30 20938126, Tel.: + 49 (0)30 20938121,
E-mail: erwin.schneider@rz.hu-berlin.de
Abbreviations: CelB
trunc
, C-terminally truncated CelB protein; CMC,
carboxy methyl cellulose; EGF, endoglucanase F; GH, glycoside
hydrolase; pNPAraf,p-nitrophenyl-a-
L
-arabinofuranoside.
Note: Nucleotide sequence data are available in the EMBL database
under the accession number AJ551527.
(Received 20 June 2003, accepted 8 July 2003)
Eur. J. Biochem. 270, 3593–3602 (2003) FEBS 2003 doi:10.1046/j.1432-1033.2003.03744.x

of their membrane lipids. Members of this genus are
acidophilic, strictly aerobic and have been described as
noncellulolytic [4]. Alicyclobacillus acidocaldarius (ATCC
27009) was first isolated from an acidic creek in Yellowstone
National Park, USA [8] and displays pH and temperature
optima of 3–4 and 60 C, respectively. Recently, we
succeeded in the cloning, purification and crystallization
of a cytoplasmic family 9 endoglucanase (CelA) from
A. acidocaldarius [9,10]. The enzyme was active against
cellobiosides, suggesting a role in degradation of imported
oligosaccharides. Here, we report the gene cloning, sequen-
cing and characterization of an extracellular endoglucanase
(CelB) from the same organism that hydrolyses carboxy
methyl cellulose (CMC), acid-swollen cellulose and oat spelt
xylan. The enzyme displays a high degree of sequence
similarity with members of GH family 51 of arabinofur-
anosidases, but completely lacks this activity. Moreover,
CelB is the first acidophilic addition to the family, exhibiting
maximal activity at pH 4 and a remarkable tolerance to pH
values ranging from 1 to 7.
Experimental procedures
Bacterial strain and culture conditions
A. acidocaldarius ATCC 27009 was grown in minimal salt
medium as described [11]. Carbon sources (at 0.2% each)
were oat spelt xylan, birchwood xylan (Roth, Germany),
starch (Sigma, Germany), sugar beet arabinan (Megazyme,
Ireland), CMC (Serva Feinbiochemica, Germany) or
glycerol. Maltose (Roth, Germany), cellobiose, glucose or
xylose (Merck, Germany) were added to a final concentra-
tion of 10 m
M
.
Cloning procedures and plasmid constructions
Restriction mapping, subcloning and Southern hybridiza-
tion were carried out using standard molecular biology
techniques according to [12]. Plasmid and phagemid
DNA was purified with Qiagen’s plasmid kit. DNA
sequencing was carried out commercially by Agowa
(Berlin, Germany) on both strands according to the
method of [13].
Chromosomal DNA from A. acidocaldarius was isolated
as described in [11]. After partial digestion of DNA with
SauIIIA, DNA fragments ranging from 8 to 12 kb were
ligated into the Zap Express vector (Stratagene, Heidelberg,
Germany), packaged using the Gigapack cloning kit
(Stratagene) and plated using Escherichia coli xl1 MRF¢
(Stratagene) as host strain according to the manufacturer’s
instructions. Screening took place by overlaying replica
plates with top agar containing 10 m
M
isopropyl thio-b-
D
-galactoside,1%CMC,250m
M
b-alanine, pH 3.5, 1 m
M
MgSO
4
,1.25m
M
CaCl
2
, 0.55% Gelrite (Merck) and
incubating overnight at 57 C. The relatively high concen-
tration of b-alanine buffer should ensure a low pH of the
top agar in order to select for acidophilic enzymes. Lysis
zones around positive plaques were identified by flooding
the plates with 0.1
M
Tris, pH 8, and staining with Congo
red according to [14]. Phagemids were derived and plated
from positive plaques according to Stratagene using the
ExAssist helper phage and the E. coli XLORL strain
(Stratagene). The resulting plasmid harboring a 6.4-kb
fragment was designated pKE25 (Fig. 1A).
Plasmid pKE2201 was constructed by ligating a PstI-
EcoRI fragment of pKE25 (Fig. 1A,B) into the expres-
sion vector pBAD/HisB (Invitrogen). The resulting ORF
(celB
trunc
) translates into a protein with six histidine residues
fused to Gly35 of the precursor. As the 3¢region of the
truncated ORF lacked a termination codon, a stop codon
provided by the pBAD/HisB vector is used. This resulted
in an extension of the protein by the sequence
PKNSKLGCFFG C-terminal of Asp-757.
Plasmid pKE25a5 was obtained by ligating a 5.7-kb KpnI
fragment that was identified by Southern hybridization of
digested chromosomal DNA with a digoxygenin-labeled
(Boehringer) XhoI-NcoIfragmentofpKE25intoplasmid
pUC18 [15] (Fig. 1A).
Plasmid pKE101, harboring the complete celB gene
was constructed by fusing the inserts from pKE25a5 and
pKE2201 via a unique KpnIsiteinpBAD/HisB.Thus,
Fig. 1. Overview of the celB region and cloning strategy, and sequence
analysis of the 5¢region of the celB gene. (A) Overview of the celB
region and cloning strategy. Shown is the celB region of the A. aci-
docaldarius chromosome (top line). Numbers indicate nucleotide
positions relative to the 5¢-SauIIIA site of the original clone (pKE25).
ORFs are represented by arrows in the direction of transcription.
Dashed arrows show ORFs neighboring celB with putative assign-
ments. The crooked arrow indicates the celB promoter detailed in B.
The thick vertical bar indicates the end of the ORF in celB
trunc
.
Restriction sites relevant to the cloning strategy are given. At the
bottom inserts of the constructed plasmids are drawn in relation to the
celB region with the restriction sites used for excision of the insert prior
to ligation in the host vector (in parentheses). The DNA fragment of
pKE25 used for Southern hybridization is marked by a black box.
(B) Sequence analysis of the 5¢region of the celB gene. Shown are the
nucleotide sequence and the corresponding amino acids. Indicated for
the nucleotide sequence are the putative )10 and )35 promoter regions
(underlined), the ribosome-binding site (double-underlined), the start
codon (boldface) and the PstI site used for subcloning (dotted line).
Indicated in the amino acid sequence are the putative cleavage site of
the signal peptidase (arrow) and the amino acid sequence found in the
N-terminus of the wild-type protein (identical positions underlined).
3594 K. Eckert and E. Schneider (Eur. J. Biochem. 270)FEBS 2003

recombinant full-length CelB has an N-terminus identical
with CelB
trunc
, but is derived from the full-length ORF with
the wild-type termination codon (see also Fig. 1A).
Computer-aided sequence analyses
Sequences were analyzed using
DNASIS
(Hitachi). The
hydropathy plot was obtained using the algorithm of Kyte
and Doolittle [16] with a window size of 50. Database
searches were conducted with
BLASTP
2.2.5 and
PSI
-
BLAST
at
NCBI [17]. Internal sequence similarities and local align-
ments between two sequences were analyzed using
PLALIGN
2.1 [18].
CLUSTALX
[19] was used for alignments and
construction of phylogenetic trees with the neighbor-joining
method. Figures were drawn with
GENEDOC
[20] and
TREE-
VIEW
[21].
Purification of wild-type CelB
A. acidocaldarius cells were grown for three days on oat
spelt xylan, reaching an OD at 650 nm of 2, harvested by
centrifugation and resuspended in the same volume of
minimal salt medium. Subsequently, cells were extracted
with Triton X-100 (0.05% final) for 30 min at 57 Cand
recentrifuged for 15 min at 20 000 g. Routinely, 450 mL of
Triton extract were adjusted to pH 6.5 by adding 10 m
M
BisTris buffer, and loaded onto a Q-Sepharose (Sigma)
column (bed volume: 25 mL) equilibrated with 10 m
M
BisTris, pH 6.5, containing 0.94 m
M
CaCl
2
,2m
M
MgSO
4
,
and 0.005% Triton X-100 (buffer A). After washing with
150 mL buffer A, elution was performed with a NaCl
gradient from 0 to 0.2
M
in 500 mL of buffer A. CelB-
containing fractions were collected, supplemented with
b-alanine buffer, pH 3.5, to a final concentration of
40 m
M
andstoredat)80 C.
Purification of recombinant CelB and CelB
trunc
E. coli strain TOP10 (Invitrogen) hosting either the
plasmid pKE101 for production of full-length CelB or
pKE2201 for CelB
trunc
was grown in LB broth, contain-
ing ampicillin (100 lgÆmL
)1
), to D
650
¼0.5. Expression
of celB and celB
trunc
was induced by addition of 0.2 and
0.02% arabinose, respectively, and growth continued for
4 h. Subsequently, cells were harvested, resuspended in
buffer B (50 m
M
sodium phosphate, pH 7, 300 m
M
NaCl, 0.1 m
M
phenylmethylsulfonyl fluoride) to a D
650
of 25. Purification of full-length CelB proceeded with
subsequent disintegration of the cells by sonication for
5 min (Sonifier II, Branson) followed by centrifugation at
130 000 gfor 1 h at 4 C. The resulting supernatant
(2 mL) was mixed with 0.5 mL Ni-NTA agarose (Qia-
gen) and Tween 20 was added to a final concentration of
0.1%. From hereon, Tween 20 and phenylmethylsulfonyl
fluoride were present in all buffers. Binding took place
for 30 min at 4 C at an imidazole concentration of
10 m
M
after which the matrix was transferred to a
column (diameter 0.5 cm) and washed with 5 mL of
buffer) B containing 10 m
M
imidazole. Elution of bound
protein was performed by raising the imidazole concen-
tration stepwise from 25 to 200 m
M
. CelB-containing
peak fractions were pooled and dialyzed overnight
(dialysis tubing type 20, 12–16 kDa cut-off, Biomol,
Germany) against buffer C (50 m
M
b-alanine, pH 3.5,
10 m
M
CaCl
2
,10m
M
MgCl
2
).
CelB
trunc
was purified by disrupting the resuspended cells
in a French press at 18 000 psi. After centrifugation 50 mL
of the resulting supernatant were diluted 1 : 1 with buffer B
and incubated with 5 mL Ni-NTA agarose for 30 min in
the presence of 10 m
M
imidazole. Then, the resin was
transferred to a column (diameter 1.5 cm), washed with
50 mL buffer B, containing 20 m
M
imidazole and protein
was eluted with 65 mL buffer B, containing 50 m
M
imidazole. Peak fractions were pooled, concentrated
10-foldwithanAmiconconcentrator(YM30membrane)
and dialyzed overnight against buffer C.
Enzyme assays
Under standard conditions enzyme activity was assayed at
a protein concentration of 1.3 lgÆmL
)1
in buffer C with
0.25% CMC for 1 h at 70 C. Subsequently, reducing sugar
content was determined according to [22]. One unit (U) is
defined as the amount of enzyme releasing 1 lmol of
reducing equivalents per minute. Xylanase activity was
measured accordingly using oat spelt xylan solubilized as
described previously [9]. In addition to the substrates used
for cultivation, linear arabinan from beet arabinan (Mega-
zyme, Ireland), avicel PH101 (Fluka, Germany), phosphoric
acid-swollen cellulose, prepared according to Wang et al.
[23] (0.25% each), and pNPAraf(Sigma, Germany)
(10 m
M
) were employed. pH stability was determined by
incubating concentrated CelB
trunc
(25 lgÆmL
)1
)in75 m
M
of
the indicated buffers, supplemented with 10 m
M
CaCl
2
and
10 m
M
MgCl
2.
After incubation overnight at 4 C, the
sample was diluted 40-fold in buffer C and activity was
assayed under standard conditions.
Thin-layer chromatography
After substrate hydrolysis in buffer C analysis of released
products was performed as described previously [9,24].
N-Terminal amino acid sequence analysis
Protein samples (40-fold concentrated Triton extract or
purified wild-type CelB) were subjected to SDS/PAGE and
stained with Serva Blue R, after which CelB was exci-
sed. Cyan bromide treatment and sequencing were
kindly performed by R. Schmid (University of Osnabru
¨ck,
Germany) as described [25,26].
Analytical methods
SDS/PAGE and staining with Serva Blue R (Serva) was
carried out as described in [11] using 10% acrylamide. Silver
staining was performed according to [27]. For activity
staining SDS gels containing either 0.2% CMC or 0.2% oat
spelt xylan were used and treated with 50 m
M
b-alanine,
pH 3.5, 0.94 m
M
CaCl
2
,2m
M
MgSO
4
according to [28].
The number of washing steps was reduced to three.
Subsequent incubation took place for 1 h at 57 C.
Immunoblot analyses were performed by transferring
proteins from SDS gels onto nitrocellulose membranes
FEBS 2003 A thermoacidophilic cellulase from Alicyclobacillus (Eur. J. Biochem. 270) 3595
using a Trans-Blot semidryapparatus (Bio-Rad) [29].
Subsequently, the membranes were incubated with a
polyclonal antiserum raised against purified wild-type CelB
(Biogenes, Germany). Antigen–antibody complexes were
visualized with peroxidase-conjugated donkey anti-rabbit
immunoglobulins using enhanced chemiluminescence
(Luminol, NEN, USA) and exposure to Hyperfilm (Amer-
sham-Buchler, Germany).
Protein concentration was determined with the Micro
BCA Protein Assay Reagent Kit (Pierce).
Results
Purification of a xylan-degrading enzyme from
A. acidocaldarius
In the initial stage of this work, we screened A. acidocalda-
rius for extracellular thermoacidophilic enzymes with poly-
saccharide-degrading activities. The organism was found to
utilize a variety of polysaccharides including xylan as sole
source of carbon and energy. However, we failed to detect
xylanase activity in the culture supernatant. Thus, assuming
a cell-associated enzyme, we succeeded in extracting xylan-
degrading activity from intact cells with Triton X-100. The
xylanase activity remained cell-bound, even after the culture
reached the stationary phase of growth. SDS/PAGE of
Triton-extracted proteins followed by silver staining
revealed about 10 major proteins with molecular masses
ranging from 30 to 100 kDa (Fig. 2A, lane 1). Zymogram
analysis demonstrated that at least five of these proteins
displayed activity towards oat spelt xylan (not shown) and
CMC (Fig. 2A, lane 2). To begin with, we concentrated our
further efforts on the 100-kDa protein. Purification of the
protein was achieved by ion exchange chromatography
using Q-Sepharose in the presence of 0.05% Triton X-100
(Fig. 3, lane 1). From a 1-L culture of A. acidocaldarius
2.0 mg of the 100-kDa protein exhibiting, on average, a
CMCase activity of 10.3 UÆmg
)1
and a xylanase activity of
0.9 UÆmg
)1
were obtained routinely.
N-Terminal sequence analysis of the protein revealed the
peptide sequence ADV(T?)STPI(A?)MEXQV, while ana-
lysis of a peptide fragment generated by cyanogen bromide
gave rise to the sequence (M)VAEL(G?)REINAY. No
homology to an entry in the database was found using
BLASTP.
The purified 100-kDa protein was injected into rabbits to
raise polyclonal antibodies. Subsequent immunoblot ana-
lysis of the Triton extract revealed that, in addition to the
100-kDa protein, two other protein bands strongly cross-
reacted with the antiserum (Fig. 2A, lane 3). This result may
imply that these proteins represent degradation products of
the 100-kDa protein. Thus, the additional bands observed
in the zymogram (Fig. 2A) are likely to represent other
enzymes that participate in the complete degradation of
CMC or xylan.
Furthermore, Western blot analysis of Triton extracts
from A. acidocaldarius cells grown on different substrates
demonstrated that, in addition to oat spelt xylan, synthesis
of the 100-kDa protein was induced by birchwood xylan,
CMC, and cellobiose, but not by glycerol, glucose, xylose,
maltose, starch or arabinan (Fig. 2B).
Cloning and sequence analysis of the 100-kDa protein
The cloning procedure with the Zap Express vector (see
Experimental procedures for details) yielded a gene bank
with 2 ·10
6
plaque-forming units (p.f.u.) with insert sizes
ranging from 3–10 kb. Screening of 45 000 plaques for
Fig. 2. Identification of CelB in Triton extract from A. acidocaldarius.
(A) Triton extract (25 lL per lane) from cells grown on oat spelt xylan
after SDS/PAGE and silver staining (lane 1), zymogram analysis with
CMC (lane 2) and Western blotting (lane 3) with antibodies raised
against wild-type CelB. (B) Western blots of Triton extracts (25 lL)
from A. acidocaldarius grown on different substrates. Lanes 1, cello-
biose; 2, starch; 3, arabinan; 4, birchwood xylan; 5, xylose; 6, CMC; 7,
glycerin;8,glucose;9,maltose.
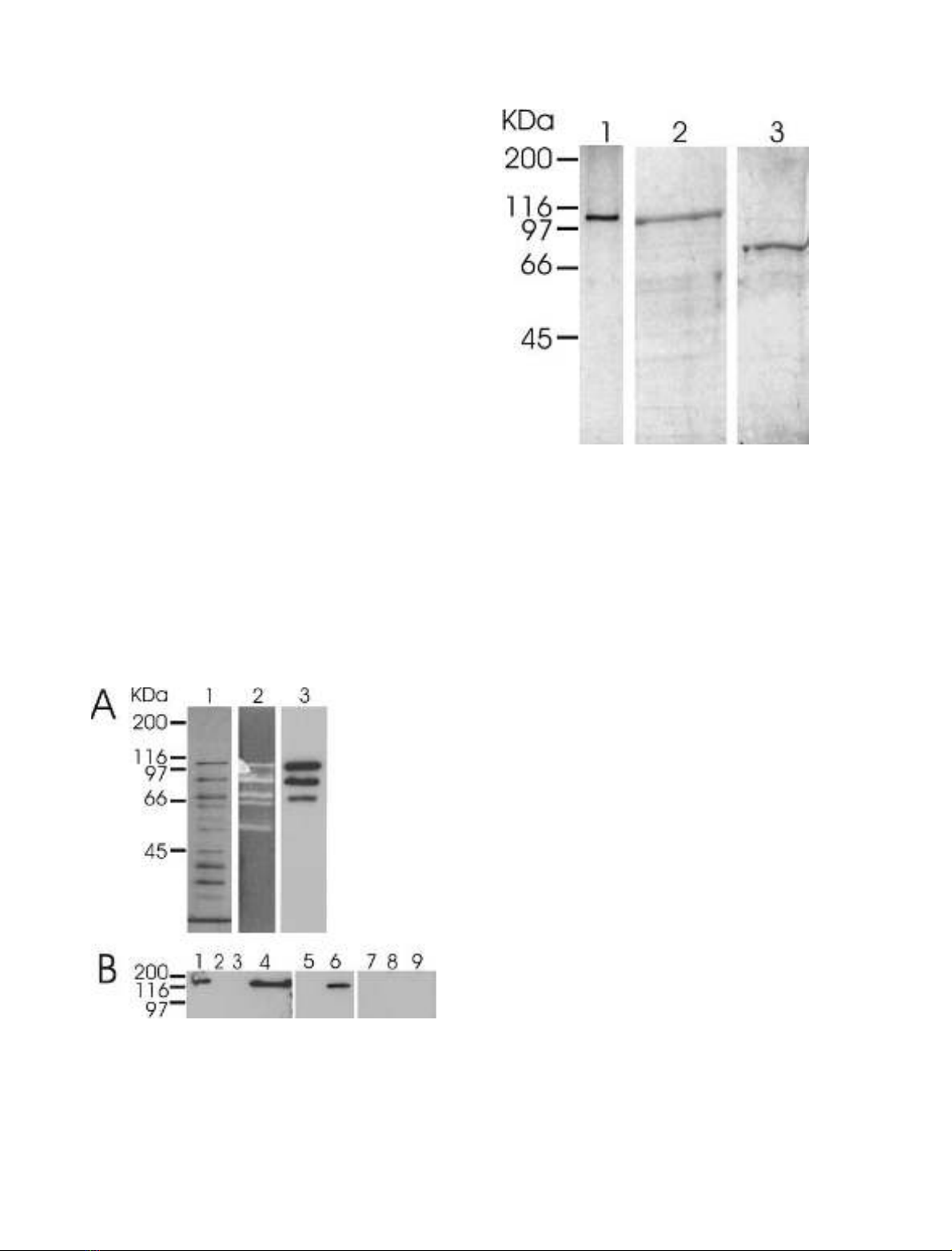
Fig. 3. SDS/PAGE of purified wild-type and recombinant forms of
CelB. Lane 1, wild-type CelB (0.2 lg), silver stained; 2, full-length
recombinant CelB (3 lg),stainedwithServaBlueR;3,recombinant
CelB
trunc
(3 lg), stained with Serva Blue R.
3596 K. Eckert and E. Schneider (Eur. J. Biochem. 270)FEBS 2003

CMC activity with the substrate overlay method and
subsequent excision resulted in five phagemids. One clone
harbored a previously described intracellular cellulase CelA
[9] as identified by Western blotting, but a second clone
reacted with antibodies raised against the wild-type 100-
kDa protein. Nucleotide sequencing revealed an incomplete
ORF which coded for a protein that displayed high
sequence similarities with endoglucanases and arabinofur-
anosidases. Eventually, screening digested chromosomal
DNA by Southern hybridization with a fragment from the
3¢end of the incomplete ORF gave rise to an overlapping
clone that contained the rest of the ORF.
The complete ORF encoded a preprotein of 959 amino
acids with a molecular mass of 100 849 kDa. A possible
start codon (TTG) with a putative ribosome-binding site
was found together with possible )10(TATAAC) and
)35(TTGACA) regions (Fig. 1B). SignalP [30] detected a
possible signal peptide whose cleavage site was located
C-terminal of amino acid Ala25 of the preprotein
(Fig. 1B). Nineteen amino acids situated C-terminally of
the cleavage site with a sequence with 79% identity to the
sequence obtained from the N-terminus of the purified
wild-type 100-kDa protein were found (Fig. 1B). More-
over, residues 485–496 of the translated ORF showed
only one substitution when compared with the internal
sequence of the 100-kDa protein (see above). In both
cases, the observed mismatches concerned only those
residues that could not unequivocally be identified by
amino acid analysis. Taken together, we conclude that the
ORF is likely to encode the 100-kDa protein that was
purified from A. acidocaldarius cells. The ORF was
designated celB.
The celB gene is flanked by two divergently transcribed
putative ORFs, encoding a LacI/GalR type transcription
regulator (152 nucleotides downstream of celB)andaLysE
type exporter (176 nucleotides upstream of celB), respect-
ively (Fig. 1A).
A database search using
BLASTP
revealed highest
sequence similarity of CelB (28% identity, 45% similarity
over a length of 410 amino acids) with endoglucanase F
(EGF) from Fibrobacter succinogenes S85 ([31], GenBank
accession number U39070) which belongs to family 51 of
glycoside hydrolases (GH51). Among the 32 other mat-
ches found, 19 were arabinofuranosidases. After three
iterations
PSI
-
BLAST
showed only three proteins not
classified as arabinofuranosidases among the top 30
matches. Sequence comparison of CelB with all members
of GH51 revealed a central catalytic domain ranging from
amino acids Thr223 to Pro702. Catalytic residues have
been inferred from sequence alignments in this family [32]
and have been experimentally confirmed [33–35]. The
conserved motif Gly Asn Glu is also present in CelB
identifying Glu366 as the acid/base catalyst. Furthermore,
Glu510 is a possible candidate for the catalytic nucleophile
residue. A phylogenetic tree constructed from the align-
ment of the catalytic domains showed that CelB forms
a distinct cluster with EGF (Fig. 4). These two are the
only enzymes characterized as endoglucanases in GH
family 51.
Adjacent to the catalytic region, a stretch of 20 amino
acids (residues Ser720–Asp739) was found with 60% of the
residues being proline, aspartate, serine or threonine, which
are typical of linker sequences [36]. This was the highest
occurrence of these amino acids in the whole sequence.
Thus, this region may function as a linker between the
catalytic domain and the C-terminal portion of the enzyme.
A database search with the N-terminal region of CelB
(residues 1–222) revealed no significant similarities to other
proteins.
Fig. 4. Phylogenetic tree of catalytic domains belonging to GH family 51.
CelB (doubly underlined) forms a distinct group with EGF (under-
lined). Also underlined is AbjA (CAA76421) from the thermophile
T. xylanilyticus.Barlength,extentofexchangeof0.1perresidue.
GenBank/GenPept accession codes are given (Agrobacterium tume-
faciens C58: AAL43920, ORF Atu3104; Arabidopsis thaliana:
AAD40132, ORF At5g26120/T1N24.13; AAF19575, ORF
At3g10740/T7M13–18; Aspergillus niger: AAC41644, arabinofurano-
sidase A; A. niger var. awamorii: IFO4033,BAB21568, ArfA; Bacil-
lus halodurans C-125: BAB05580, ORF AbfA (BH1861); BAB05593,
ORF Xsa (BH1874); Bacillus subtilis ssp. subtilis str. 168: CAA61937,
arabinofuranosidase 1; CAA99576, arabinofuranosidase 2; Bactero-
ides ovatus: AAA50391, arabinosidase 1; AAA50393, arabinosidase 2;
Bifidobacterium longum NCC2705: AAN24035, BL0181; AAN24368,
AbfA1; AAN24945, BL1138; AAN24971, AbfA2; AAN25400, AbfA3;
Caulobacter crescentus CB15: AAK23403, ORF CC1422; Cellvibrio
japonicus: AAK84947, arabinofuranosidase; Clostridium acetobutyl-
icum ATCC 824: AAK81366, ORF CAC3436; Clostridium sterco-
rarium: AAC28125, arabinofuranosidase; Cytophaga xylanolytica:
AAC38456, arabinofuranosidase I; AAC38457, arabinofuranosidase
II; F. succinogenes S85: AAC45377, EGF; G. stearothermophilus
T-6: AAD45520, abfA; Hordeum vulgare: AAK21879, AXAH-I;
AAK21880, AXAH-II; Mesorhizobium loti MAFF303099: NP 104667,
Mll3591; Oryza sativa: BAC10349, OJ1200 C08.20; CAD39869,
OSJNBb0058J09.6; Sinorhizobium meliloti 1021: CAC49446, AbfA;
Streptomyces chartreusis: BAA90771, arabinofuranosidase I; Strepto-
myces coelicolor A3(2): CAA20794, ORF SCI35.05c; CAB86096,
AbfA; Streptomyces lividans 66: AAA61708, AbfA; T. xylanilyticus
D3: CAA76421, AbjA; Thermotoga maritima: AAD35369, ORF
TM0281).
FEBS 2003 A thermoacidophilic cellulase from Alicyclobacillus (Eur. J. Biochem. 270) 3597


























%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)