
Salt-induced formation of the A-state of
ferricytochrome c– effect of the anion charge
on protein structure
Federica Sinibaldi, Maria C. Piro, Massimo Coletta and Roberto Santucci
Dipartimento di Medicina Sperimentale e Scienze Biochimiche, Universita
`di Roma ‘Tor Vergata’, Italy
Formation of the unique, native structure of a protein
occurs through well-defined folding pathways involving
a limited number of intermediate species. In recent
years, a large body of kinetic and equilibrium studies
has provided extensive information on the folding
pathway of proteins and led to the characterization of
intermediate states, thus contributing to our under-
standing of the protein-folding mechanism [1–9].
The non-native compact state of equine cyto-
chrome cstabilized by salts in an acidic environment
(pH 2.0–2.2), called the A-state, is thought to be a
suitable model for the molten globule of cytochrome c;
it possesses a native-like a-helix conformation but a
fluctuating tertiary structure [10–14]. With respect to
the native protein, in the A-state some interior hydro-
phobic residues become exposed to the solvent [15],
the W59-one-heme-propionate hydrogen bond is
impaired (although the tryptophan remains within a
hydrophobic environment) [14], and the heme–poly-
peptide chain interaction is reduced. Also, the hydro-
phobic core (which is composed of the two major
helices and the heme group) is preserved in the A-state,
Keywords
A-state; cytochrome c; fast kinetics; folding;
site-directed mutagenesis
Correspondence
R. Santucci, Dipartimento di Medicina
Sperimentale e Scienze Biochimiche,
Universita
`di Roma ‘Tor Vergata’,
V. Montpellier 1, 00133 Roma, Italy
Fax: +39 06 72596353
Tel: +39 06 72596364
E-mail: santucci@med.uniroma2.it
(Received 1 August 2006, revised 28 sep-
tember 2006, accepted 5 October 2006)
doi:10.1111/j.1742-4658.2006.05527.x
Structural information on partially folded forms is important for a deeper
understanding of the folding mechanism(s) and the factors affecting protein
stabilization. The non-native compact state of equine cytochrome cstabil-
ized by salts in an acidic environment (pH 2.0–2.2), called the A-state, is
considered a suitable model for the molten globule of cytochrome c,asit
possesses a native-like a-helix conformation but a fluctuating tertiary struc-
ture. In this article, we extend our knowledge on anion-induced protein sta-
bilization by determining the effect of anions carrying a double negative
charge; unlike monovalent anions (which are thought to exert an ‘ionic
atmosphere’ effect on the macromolecule), divalent anions are thought to
bind to the protein at specific surface sites. Our data indicate that divalent
anions, in comparison to monovalent ions, have a greater tendency to sta-
bilize the native-like M–Fe(III)–H coordinated state of the protein. The
possibility that divalent anions may bind to the protein at the same sites
previously identified for polyvalent anions was evaluated. To investigate
this issue, the behavior of the K88E, K88E ⁄T89K and K13N mutants was
investigated. The data obtained indicate that the mutated residues, which
contribute to form the binding sites of polyanions, are important for stabil-
ization of the native conformation; the mutants investigated, in fact, all
show an increased amount of the misligated H–Fe(III)–H state and, with
respect to wild-type cytochrome c, appear to be less sensitive to the pres-
ence of the anion. These residues also modulate the conformation of unfol-
ded cytochrome c, influencing its spin state and the coordination to the
prosthetic group.
Abbreviation
CT, charge transfer.
FEBS Journal 273 (2006) 5347–5357 ª2006 The Authors Journal compilation ª2006 FEBS 5347
stabilized by nonbonded interactions [12,16], whereas
the loop regions appear to be fluctuating and partly
disordered [12]. The A-state is promptly achieved at
pH around 2.2 upon addition of a salt to an aqueous
HCl solution containing denatured cytochrome c; this
has been ascribed to a screening action of the anions,
which stabilize the compact form by binding to the
positively charged groups on the protein surface [11].
Recently, we investigated the role played by mono-
valent anions in promoting the transition from the
acid-denatured protein to the A-state [17,18]. Our
results showed that the salt-induced A-state of ferri-
cytochrome cis characterized by a variety of high-
spin and low-spin states (where ‘high’ and ‘low’ stand
for the S¼5⁄2 and S¼1⁄2 spin states of the heme
iron, respectively) in equilibrium; in particular (at
least), two distinct low-spin species, differing in their
axial ligation to the metal, coexist in solution: a form
with the native M–Fe(III)–H coordination, and a bis-
histidine coordinated species. The equilibrium between
these two low-spin forms, here indicated as M-Fe
(III)-H ! H-Fe(III)-H is strongly influenced by the
type of anion in solution [17,18].
Because structural information on partially folded
forms is important for a deeper understanding of the
folding mechanism(s) and the factors affecting protein
stabilization, in this article we extend our knowledge
on anion–protein interactions by determining the effect
on the protein produced by anions carrying a double
negative charge. This is an interesting point to investi-
gate, because, unlike monovalent anions (which are
thought to exert an ‘ionic atmosphere’ effect on the
macromolecule), divalent anions (as well as polyvalent
groups, such as polyphosphates [19,20]) are supposed
to bind to the protein at specific surface sites [21,22].
Results
Horse ferricytochrome c
CD measurements
Far-UV CD (200–250 nm) is a probe for the formation
of the A-state from acid-denatured cytochrome c,as
the A-state possesses a native-like a-helix structure
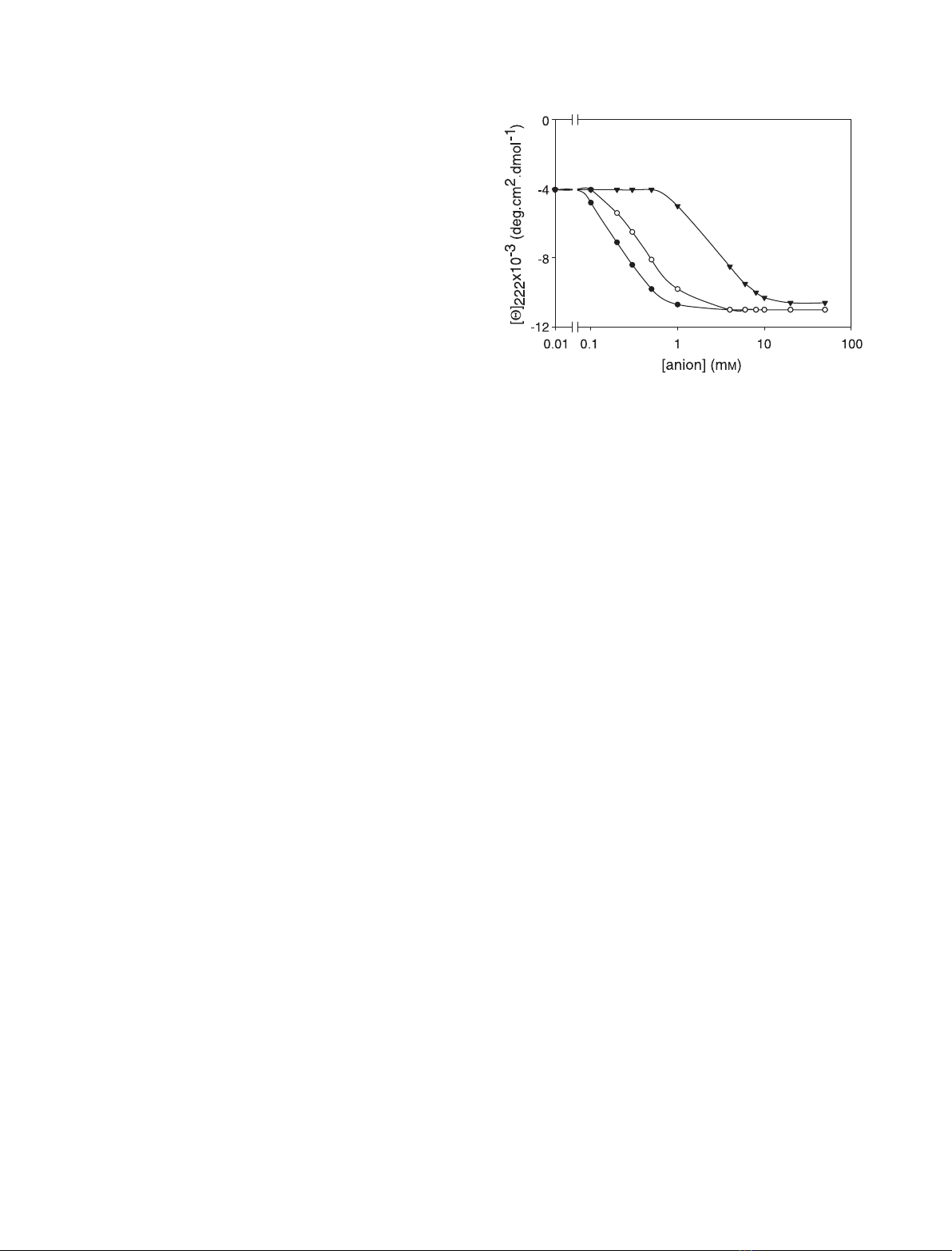
[11,17]. Figure 1 shows the gradual recovery of the
ordered secondary structure in acid-denatured ferricyt-
ochrome cupon addition of increasing amounts of
sulfate and selenate; the divalent anions stabilize the
A-state at significantly lower concentrations than those
needed for stabilization by monovalent ions [17]. As
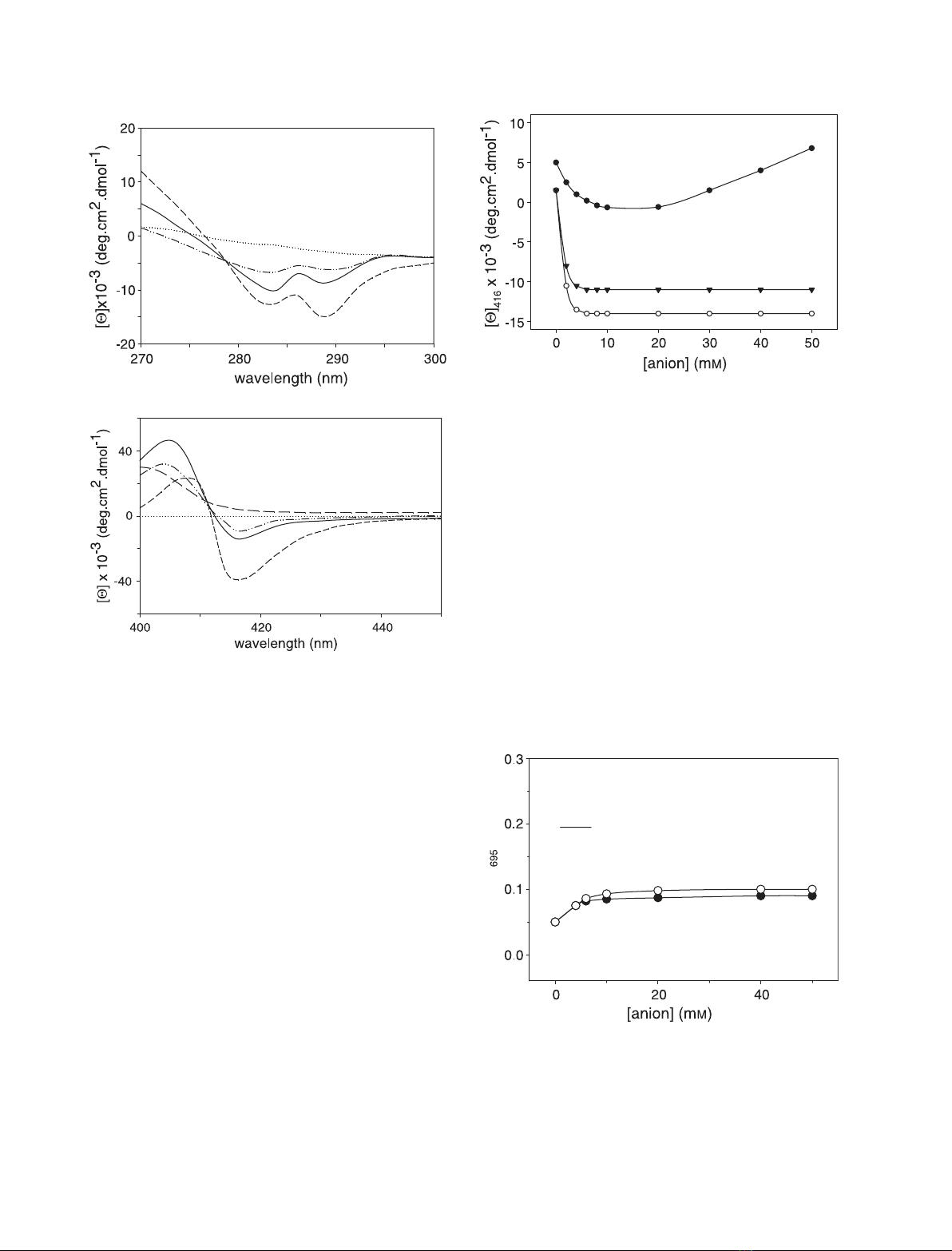
shown in Fig. 2, the A-state tertiary conformation is
less packed than that of the native form; the protein
displays a weaker near-UV CD spectrum (Fig. 2A),
consistent with a perturbed W59 microenvironment,
and a weaker Soret CD spectrum (Fig. 2B). In this last
case, the decreased intensity of the 416 nm Cotton
effect is indicative of a perturbed heme pocket region,
as the 416 nm dichroic band is considered to be diag-
nostic for the Met80–Fe(III) coordination in native
cytochrome c[23,24]). As the M–Fe(III)–H coordina-
ted species alone contributes to the dichroic signal, a
significant population of macromolecules is expected
to lack M80 coordination to Fe (III) in the A-state
(it must be noted, however, that the signal is stronger
than that recorded in the presence of monovalent ani-
ons [17,18]). The intensity of the 416 nm dichroic band
is 35% that of the native state, consistent with het-
erogeneity of the A-state. On the basis of earlier data
(relative to monovalent anions) [18], a mixture between
Met80–Fe(III)–His18 coordinated species and X-Fe-
His18 miscoordinated species (where X represents the
endogeneous ligand coordinated to the metal in place
of Met80) is expected in solution. Under the condi-
tions investigated, a histidine (His26 or His33) is
expected to be the best candidate for ligand X (the
other likely candidates, i.e. the lysines, are fully proto-
nated at pH 2.2) [18].
The heterogeneous character of the A-state promp-
ted us to investigate the effect of sulfate and selenate
on the heme pocket conformation. As shown in Fig. 3,
the 416 nm dichroic band gradually increases (towards
negative ellipticity values) with anion concentration,
up to 4 mmanion; it then remains unchanged (up to
40 mmanion). This behavior markedly differs from
that displayed by the protein in the presence of
Fig. 1. Sulfate-induced (d) and selenate-induced (s) conformational
transition of acid-denatured cytochrome cto the A-state, as meas-
ured by the ellipticity at 222 nm. Experimental conditions: aqueous
HCl, pH 2.2; temperature 25 C. The transition in perchlorate (.)is
shown for comparison.
Anion-modulated structure of cyt cA-state F. Sinibaldi et al.
5348 FEBS Journal 273 (2006) 5347–5357 ª2006 The Authors Journal compilation ª2006 FEBS
monovalent anions (the effect of perchlorate is illustra-
ted in Fig. 3 for comparative purposes). The changes
in the Cotton effect strength observed at high mono-
valent anion concentrations have been attributed to a
shift of the M-Fe(III)-H ! H-Fe(III)-H equilibrium
towards formation of the bis-H species [18]. Thus,
the data in Fig. 2 indicate that divalent anions have
a stronger tendency to stabilize the (native-like)
M–Fe(III)–H coordinated form.
Unfolded macromolecules and peptides attain a
degree of structure at temperatures lower than room
temperature. We have recently shown that the A-state
induced by monovalent anions displays a temperature-
dependent 416 nm Cotton effect (temperature range:
25 Cto2C) [17]. In the present study, the investiga-
tion, extended to divalent anions, confirms that the
native M–Fe(III)–H bond (indicative of a more struc-
tured conformation) is stabilized by low temperature
(data not shown), indicating that protein flexibility hin-
ders methionine coordination to the heme iron [25].
Electronic absorption
The 695 nm absorption band is considered to be diag-
nostic for the M80–Fe(III) axial bond in native cyto-
chrome c[26]. Figure 4 shows the effect of sulfate and
selenate on acid-denatured cytochrome c, investigated
Fig. 3. Effect of sulfate (s) and selenate (.) concentration on the
heme pocket environment [and on the strength of the Met80–
Fe(III) axial bond] of the salt-induced A-state of cytochrome c,as
observed from changes induced in the 416 nm Cotton effect. The
effect induced by the monovalent anion perchlorate (d) is reported
for comparison. Other experimental conditions were as described
in the legend to Fig. 1.
A
Fig. 4. Absorbance at 695 nm of acid-denatured cytochrome cin
the presence of increasing sulfate (s) and selenate (d) concentra-
tions. The optical absorbance of native cytochrome c(—) at pH 7.0
is shown for comparison. Protein concentration: 0.25 mM. Other
experimental conditions were as described in the legend to Fig. 1.
A
B
Fig. 2. Near-UV (A) and Soret (B) CD spectra of acid-denatured
cytochrome cin the presence of 0.02 Msulfate (—) and 0.02 Msel-
enate (— ÆÆ—ÆÆ). The spectra of the native (-Æ-Æ-) and of the dena-
tured (ÆÆÆÆ) protein are shown for comparison. Protein concentration:
10 lM. Other experimental conditions were as described in the
legend to Fig. 1.
F. Sinibaldi et al.Anion-modulated structure of cyt cA-state
FEBS Journal 273 (2006) 5347–5357 ª2006 The Authors Journal compilation ª2006 FEBS 5349
by following the changes in the 695 nm absorbance
band. It appears clear that both anions favor protein
collapse into a compact form, and induce formation of
a consistent population of macromolecules (35% in
sulfate, 28% in selenate) with native M–Fe(III)–H
coordination. These data are in excellent agreement
with CD measurements and provide independent evi-
dence for heterogeneity of the A-state.
A-state stability
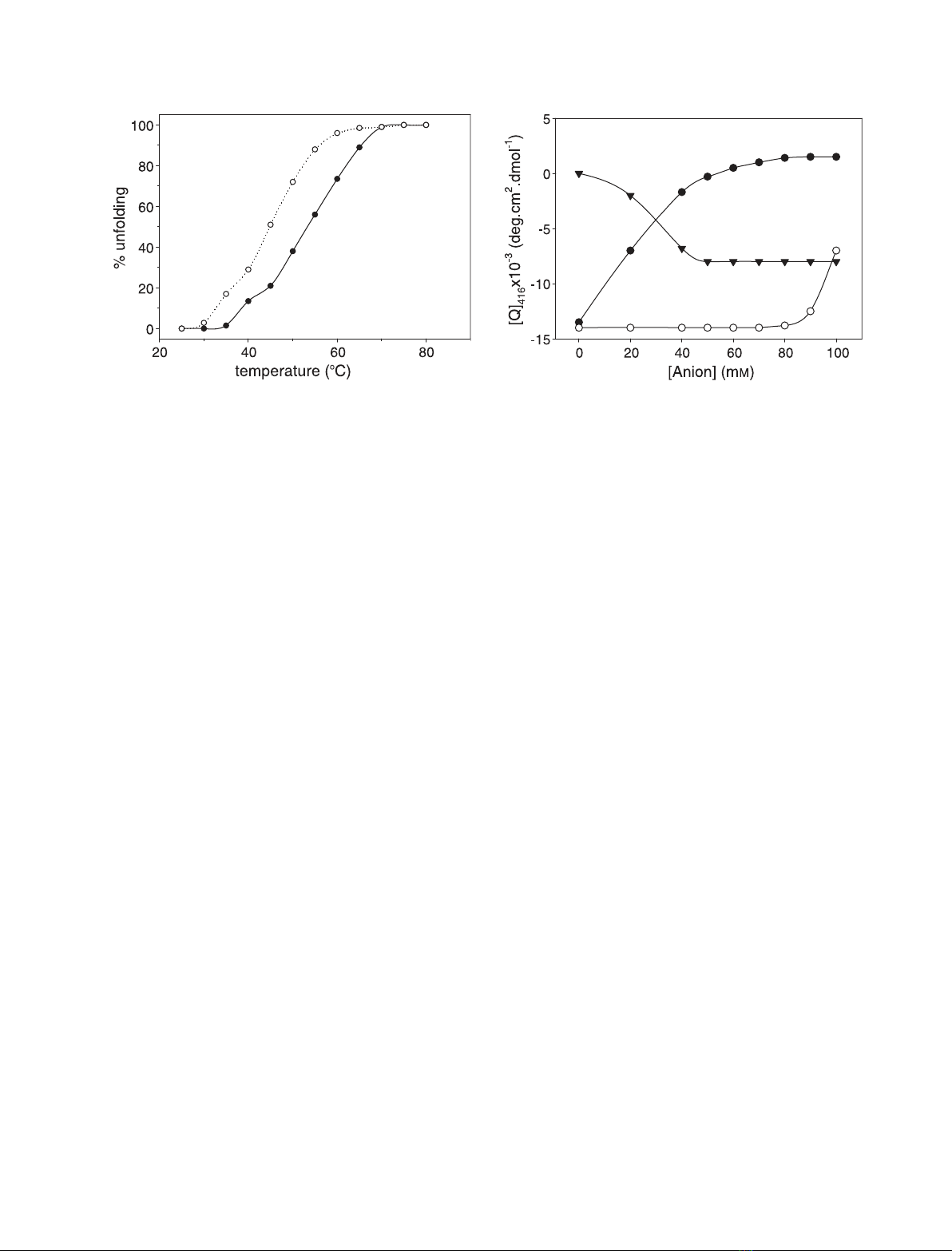
Figure 5 shows the thermal denaturation profiles of
the A-state of cytochrome c, as obtained from ellip-
ticity values at 222 nm. As previously observed for
mononvalent anions [18], the shape of the unfolding
profiles features a multiple state transition, as (at least)
three distinct thermodynamic states are detected. The
profiles clearly show that protein stability strongly
depends on anion concentration; this highlights the
primary role played by the anion–protein interactions
in A-state stabilization.
Competition among anions
To better define the effect produced by monovalent
anions on the sulfate-induced A-state of cytochrome c,
we monitored the changes in the 416 nm Cotton effect
induced by increasing amounts of perchlorate and
chloride. As shown in Fig. 6, addition of monovalent
anions alters the 416 nm dichroic band; this suggests
competition between monovalent and divalent anions
for binding to the protein. In particular, both perchlor-
ate and Cl
–
shift the M-Fe(III)-H ! H-Fe(III)-H
equilibrium towards the bis-H species, and destabilize
the M–Fe(III)–H coordinated form. The reduced effect
of Cl
–
reflects the different affinities of the two anions
for the protein [11,17].
We also monitored the effect of sulfate on the
perchlorate-induced A-state. As shown in Fig. 6, addi-
tion of sulfate strengthens the 416 nm dichroic band,
which confirms that divalent anions have a greater
tendency to stabilize the M80–Fe(III)–H18 coordinated
form. On the whole, these data support competitive
anion binding to the protein, and the idea that mono-
valent and divalent anions tend to stabilize differently
structured A-states.
Horse ferricytochrome cvariants
Anions carrying multiple negative charges bind to spe-
cific sites of horse cytochrome c[19,27]. To determine
whether divalent anions bind to the same sites, we
introduced some mutations within the site-containing
regions of the macromolecule, with the aim of defining
the role played by single residues in modulating pro-
tein affinity for divalent anions. On the basis of earlier
work [19,27], the sites under consideration were:
(a) the site encompassing residues K87, K88, and R91,
located in the C-terminal a-helix segment, indicated
here as site 1; and (b) the site encompassing residues
K86, K87, and K13, located at the interface between
the N-terminal and the C-terminal a-helices, indicated
Fig. 5. Thermal stability of the A-state of cytochrome cas a func-
tion of sulfate concentration. Sulfate concentration: s,10mM;d,
40 mM. The experimental points refer to ellipticity values at
222 nm. Other experimental conditions were as described in the
legend to Fig. 1.
Fig. 6. Effect of perchlorate (d) and chloride (s) concentration on
the heme pocket environment of the sulfate-induced A-state of
cytochrome c, as observed from changes induced in the 416 nm
Cotton effect (sulfate concentration: 50 mM). The effect of sulfate
(.) concentration on the perchlorate-induced A-state is also illustra-
ted (perchlorate concentration: 50 mM). Other experimental condi-
tions were as described in the legend to Fig. 1.
Anion-modulated structure of cyt cA-state F. Sinibaldi et al.
5350 FEBS Journal 273 (2006) 5347–5357 ª2006 The Authors Journal compilation ª2006 FEBS
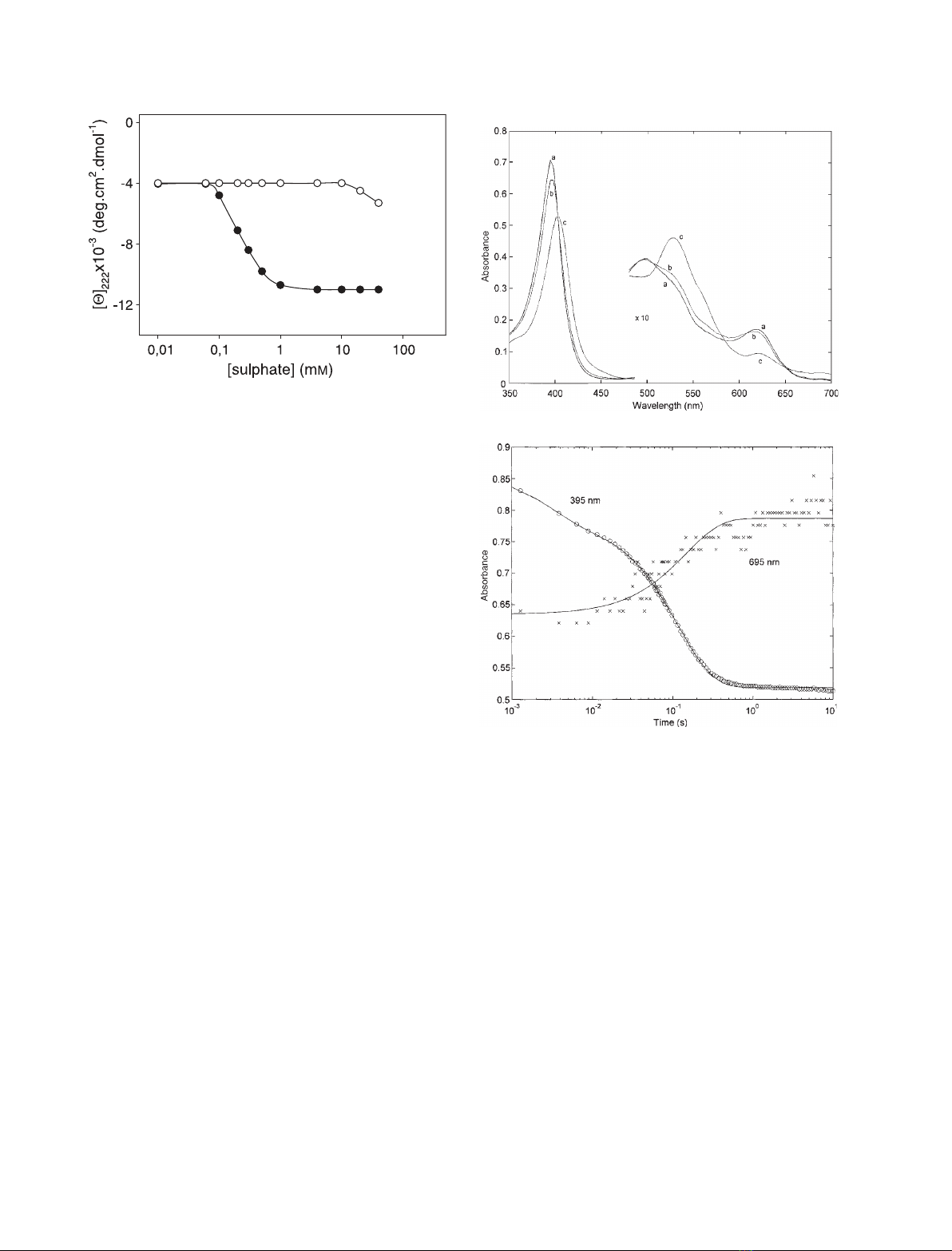
here as site 2. The residues under investigation were
substituted with residues located at the same position
in yeast iso-1-cytochrome c; as illustrated in Fig. 7,
horse and yeast cytochrome cshow very different
affinities (considered here as a nonspecific indicator of
the binding effect, not as a direct measure of anion
binding to the protein) for anions.
CD and absorption measurements
In site 1, the K88E mutation introduces an acidic resi-
due (E88, present in yeast [28]) in place of a lysine,
whereas in site 2, the K13N mutation introduces an
asparagine in place of a lysine. This provides the
opportunity to evaluate the contribution of K88 and
K13 to protein stabilization in the reaction with sul-
fate. The far-UV and Soret CD spectra of the two
mutants (not shown) reveal that the two variants and
the wild-type protein are equally influenced by sulfate.
Similar results were obtained when we investigated the
spectroscopic properties of the K88E ⁄T89K double
mutant, which, with respect to the K88E mutant, pos-
sesses a sequence closer to the corresponding sequence
in yeast iso-1-cytochrome c. A 40 mmsulfate concen-
tration induced, in all the variants investigated, native-
like a-helix content and formation of the 416 nm
Cotton effect with a strength comparable (although
not identical) to that of the wild-type protein. This
excludes the possibility that K88, T89 and K13 modu-
late horse cytochrome caffinity for anions. Also, the
mutant’s stability is not dissimilar to that of the wild-
type protein, as indicated by thermal denaturation
studies (data not shown).
Fast kinetic measurements
The 350–700 nm absorption spectrum of acid-dena-
tured cytochrome c(spectrum a of Fig. 8A) displays
an absorption maximum around 395 nm in the Soret
region, and a maximum at 497 nm, a shoulder at
528 nm and a charge transfer (CT) at 618 nm in the
visible region. The spectral changes detected at pH 2.2
Fig. 7. Sulfate-induced conformational transition of acid-denatured
horse ferricytochrome c(d) and yeast iso-1-ferricytochrome c(s)
to the A-state, as measured by the ellipticity at 222 nm. Experimen-
tal conditions: aqueous HCl, pH 2.2; temperature 25 C.
A
B
Fig. 8. (A) Absorption spectra of ferricytochrome cbefore (spec-
trum a) and after 40 ms (spectrum b) and 5 s (spectrum c) of mix-
ing with 40 mMsulfate. Absorption spectra in the visible range are
a 10-fold magnification of original spectra. (B) Kinetic progress
curves of wild-type cytochrome cafter mixing with 40 mMsulfate
at 395 nm and at 695 nm, as indicated. The progress curve at
695 nm has been magnified in order to compare its signal time
evolution with that at 395 nm. The solid lines are the least-squares
nonlinear fitting of the kinetic progress curve according to Eqn (1),
with n¼2 and with the following rate constants: k
1
¼350 ±
40 s
)1
and k
2
¼8.4 ± 0.9 s
)1
at 395 nm, and k¼7.7 ± 0.7 s
)1
at
695 nm.
F. Sinibaldi et al.Anion-modulated structure of cyt cA-state
FEBS Journal 273 (2006) 5347–5357 ª2006 The Authors Journal compilation ª2006 FEBS 5351










![Giáo trình Kỹ thuật chung về ô tô (Nghề: Công nghệ ô tô) - Trường Cao đẳng Bách Khoa Tây Nguyên [Mới Nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20251212/laphong0906/135x160/39771779074470.jpg)
![Giáo trình Kỹ thuật chung về ô tô và công nghệ sửa chữa (Nghề: Công nghệ ô tô) - Trường Cao đẳng Bách Khoa Tây Nguyên [Mới Nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20251212/laphong0906/135x160/23311779074471.jpg)



![Giáo trình Hệ thống nhiên liệu động cơ ô tô (CĐ) - Trường Cao đẳng Công nghiệp Thanh Hóa [Ngành Công nghệ ô tô]](https://cdn.tailieu.vn/images/document/thumbnail/2026/20260511/hoatrami2026/135x160/37511778728704.jpg)
![Giáo trình Hệ thống phanh ABS Công nghệ ô tô (CĐ) - Trường Cao đẳng Công nghiệp Thanh Hóa [Mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2026/20260511/hoatrami2026/135x160/1901778728704.jpg)





![Giáo trình Hệ thống điện thân xe ô tô (CĐ) - Trường Cao đẳng Công nghiệp Thanh Hóa [Mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2026/20260511/hoatrami2026/135x160/611778728708.jpg)


%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)