
Mä liãn kãút - Mä phäi
MÔ LIÊN KẾT
Mục tiêu học tập
1. Trình bày được cấu trúc và chức năng mô liên kết.
2. Phân loại được mô liên kết.
3. Mô tả được cấu trúc các sợi liên kết và các tế bào liên kết.
Mô liên kết là tập hợp những tế bào có nguồn gốc trung bì, giữ chức năng bảo vệ,
nâng đỡ làm sườn cấu tạo cho cơ thể và cơ quan.
Mô liên kết hiện diện ở khắp các cơ quan, giúp cơ thể thể hiện tính thống nhất về cấu
tạo và chức năng. Cấu tạo của mô liên kết gồm 3 thành phần chính:
1. Chất căn bản.
2. Những phân tử sợi.
3. Những tế bào liên kết.
* Phân loại: căn cứ vào tính chất của chất căn bản, người ta chia mô liên kết ra làm 3
loại:
1. Mô liên kết chính thức.
2. Mô sụn.
3. Mô xương.
MÔ LIÊN KẾT CHÍNH THỨC
I. CHẤT CĂN BẢN
Chất căn bản mô liên kết chính thức là một chất vô định hình, đồng nhất, trong suốt,
làm nền cho tế bào với các phân tử sợi, có tính nhờn với hàm lượng nước và chất điện giải
tương đương với máu, được hình thành bởi 2 loại protein chính: glycoaminoglycans và
glycoprotein cấu trúc.
1.Glycosaminoglycans
Là những chuỗi Polysaccharide được tạo với sự đa trùng hợp của những đơn vị
disaccharide gắn với acid uronic và nhóm hexosamine, những nhóm đa đường này thường gắn
với protein bởi những nối đồng hoá trị (covalent) để tạo những phân tử proteoglycan, các
protein hoà tan này thường là dermatan sulfate, chondroitin sulfate, keratan sulfate, heparan
sulfate.
- Dermatan sulfate phần lớn ở da, gân, dây chằng, sụn xơ, tất cả cấu trúc này
chứa collagene type I.
- Chondroitin sulfate có nhiều ở sụn trong và sụn đàn hồi.
- Heparan sulfate có khuynh hướng kết hợp với sợi võng và Collagene type III.
Những proteoglycan này làm cho chất căn bản ở trạng thái nửa sol nửa gel.
2. Glycoprotein cấu trúc: những protein này là:
8
Mä liãn kãút - Mä phäi
- Fibronectin: là 1 glycoprotein được tổng hợp từ tế bào sợi và tế bào biểu mô. Những
phân tử protein này giúp cho sự liên kết giữa tế bào, sợi collagene và các nhóm glycosamine,
sự liên kết này tác động đến tính liên kết của các tế bào và tính di chuyển cuả nó.
Tế bào ung thư là những tế bào không tạo ra fibronectin phần nào giải thích tính xâm
nhập và phá huỷ màng đáy của chúng.
- Laminin: là glycoprotein, 1 đại phân tử glycoprotein chứa ít nhất 1 chuỗi
polypeptide, chúng được tìm thấy ở màng đáy giúp cho sự gắn kết của biểu mô với collagene
type IV của màng đáy.
- Chondroitin có ở sụn giúp cho sự liên kết giữa tế bào sụn và collagene type II.
II. NHỮNG PHẦN TỬ SỢI
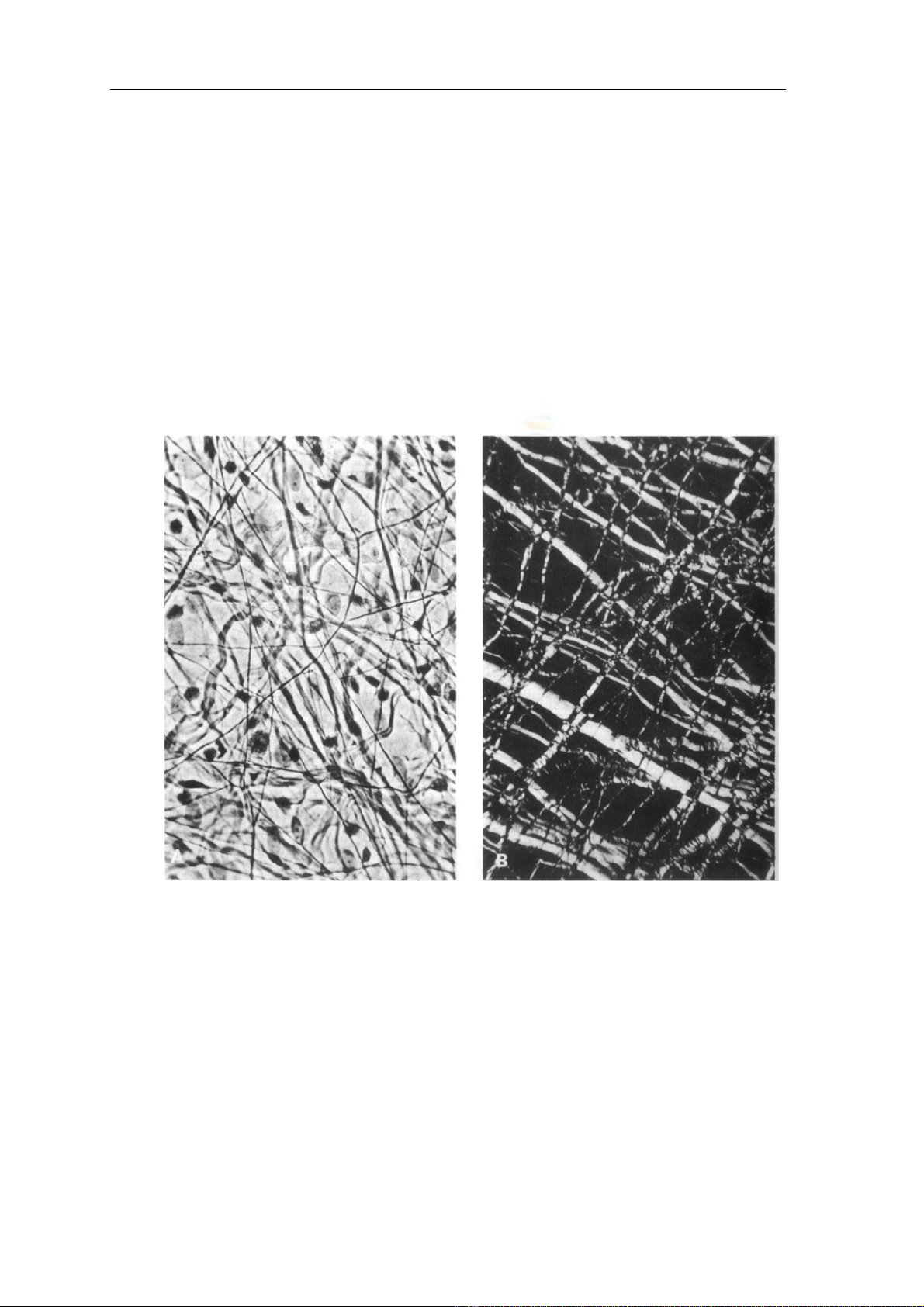
có 3 loại sợi: (Hình 1)
- Sợi collagene, sợi đàn hồi, sợi võng.
Hình 1a: Sợi tạo keo và sợi đàn hồi nhuộm Hình 1b: Sợi tạo keo và sợi đàn hồi
bằng phương pháp Weigert x 200 hiển vi phân cực
1. Sợi collagene:
Collagene là 1 loại sợi bắt màu dễ dàng với nhiều loại thuốc nhuộm dành cho hiển vi
quang học, hình thái của nó rất biến thiên tuỳ theo mô và cơ quan. Chúng phân bố dưới dạng
những sợi mảnh ở các lớp đệm (lammina propria) hoặc mô liên kết lỏng lẻo (loose connective
tissue), dày đặc dưới dạng bó sợi ở gân, dây chằng, dạng lá ở mô liên kết dưới da, những sợi cực
mảnh dàn thành tấm ở giác mạc mắt.
Ðúng tính chất của sợi là không màu nhưng vì sự sắp xếp cuả chúng cho nên gần dây
chằng có màu trắng ngà trong lúc giác mạc mắt trong suốt.Dưới hiển vi điện tử,sợi xuất hiện
dưới dạng những sợi nhỏ hợp nhau thành bó, với những băng sáng và băng tối chạy ngang,
đều đặn một cách chu kỳ, chu kỳ là 640( (Hình 2, 4).
9
Mä liãn kãút - Mä phäi
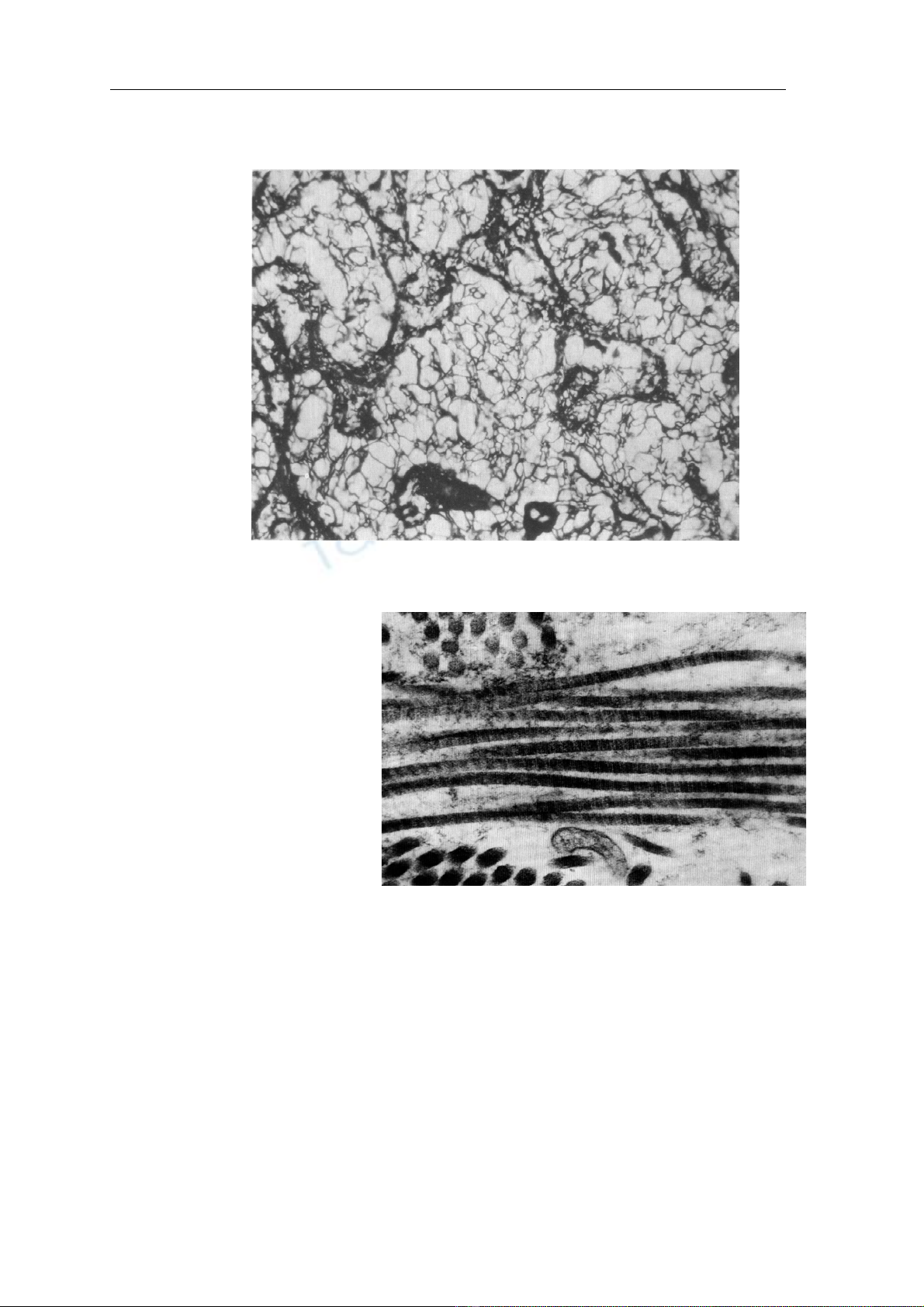
H ình 1 C:
S ợi v õng nhu ộm b
ằng ph ư ơng ph áp nhu ộm ng ấm b ạc x 200
Lúc đầu người ta không biết vì sao sợi lại có hình ảnh này, cho mãi đến năm 1950,
Groos, Schmit và Highberger mới
tìm cách tách các protein từ gian
bào chất cuả mô liên kết đang
phát triển (non) một loại protein
hình gậy có chiều dài chừng
30nm, đường kính 1,4 nm. Protein
này hoà tan trong nước muối sinh
lý ở nhiệt độ lạnh và chúng có
khuynh hướng kết hợp thành sợi ở
nhiệt độ của cơ thể, các sợi này có
hình ảnh rất giống sợi collagene
khi quan sát bằng hiển vi điện tử.
Hình 2: Sợi tạo keo hiển vi điện tử
x 100.000
Hodge và Petruska đã giải thích sự hình thành của sợi collagene một cách đầy đủ nhất,
các protein hình gậy ở trên chính là tropocollagene - một đơn vị protein cơ bản để tạo nên sợi
collagene, trong gian bào chất các tropocollagene sắp xếp theo một trình tự nghiêm ngặt để
tạo nên sợi collagene, quá trình này thường được mệnh danh là đa trùng hợp. Những protein
này sắp xếp song song, những sợi tropocollagene ở cùng một hàng cách nhau khoảng 0,6D,
sợi trên và sợi dưới chênh nhau 0,4D, chiều dài tropocollagene được tính bằng 4,4D, D =
67nm. Chính sự sắp xếp này đã tạo nên các ô lỗ lưới. Khi sử dụng osmium để cố định đồng
thời cũng là thuốc "nhuộm" trong kỹ thuật hiển vi điện tử, các muối osmium đã bị tẩm vào
các ô này, do đó trên hiển vi điện tử sợi có band sáng và band tối có chu kỳ.
Tropocollagene là một protein phức tạp được hình thành do s ự xoắn lại của 3 sợi
10
Mä liãn kãút - Mä phäi
polypeptide dưới dạng (helic, mỗi sợi polypeptide được gọi là: sợi (polypeptide. Sợi
(collagene là một trong những polypeptide dài nhất đã được biết có chừng 1050 acide amine,
trọng lượng phân tử
100000.
Ngày nay khi đề
cập đến (helic hai sợi
(double (helic) người ta
thường nghĩ ngay đến
cấu trúc của DNA -
trung tâm điều khiển
hoạt động của sinh vật.
Song ở sinh vật còn có
những cấu trúc (helic 2
sợi khác giữ chức vụ ít
quan trọng hơn DNA
nhưng góp phần rất lớn
trong hoạt động của sự
sống như cấu trúc của
protein 2 sợi (helic của
myosin đã quyết định
cho cơ chế co và dãn cơ.
Một sợi khác và
có lẽ là duy nhất có cấu
trúc 3 sợi (helic, đó là
tropocollagene ít được
để ý nhất.
Sợi ( hình thành
do sự cắt bớt các đoạn
peptide của tiền sợi ( về
phía 2 đầu. Ở 2 đầu sợi
có dạng thẳng và được
gọi là peptide xa
(telopeptide), acide
amine chiếm ưu thế
trong peptide xa là lysine
và hydroxylysine, hydro
hoá lysine để biến thành
hydroxy lysine cần
enzym đặc hiệu .
Phía amino tận của tiền
chuỗi có 1 đoạn
polypeptide rất ngắn, rất
kị nước, gọi là chuỗi tín
hiệu. Ngày nay người ta
cho rằng như mọi loại
protein khác thường kèm
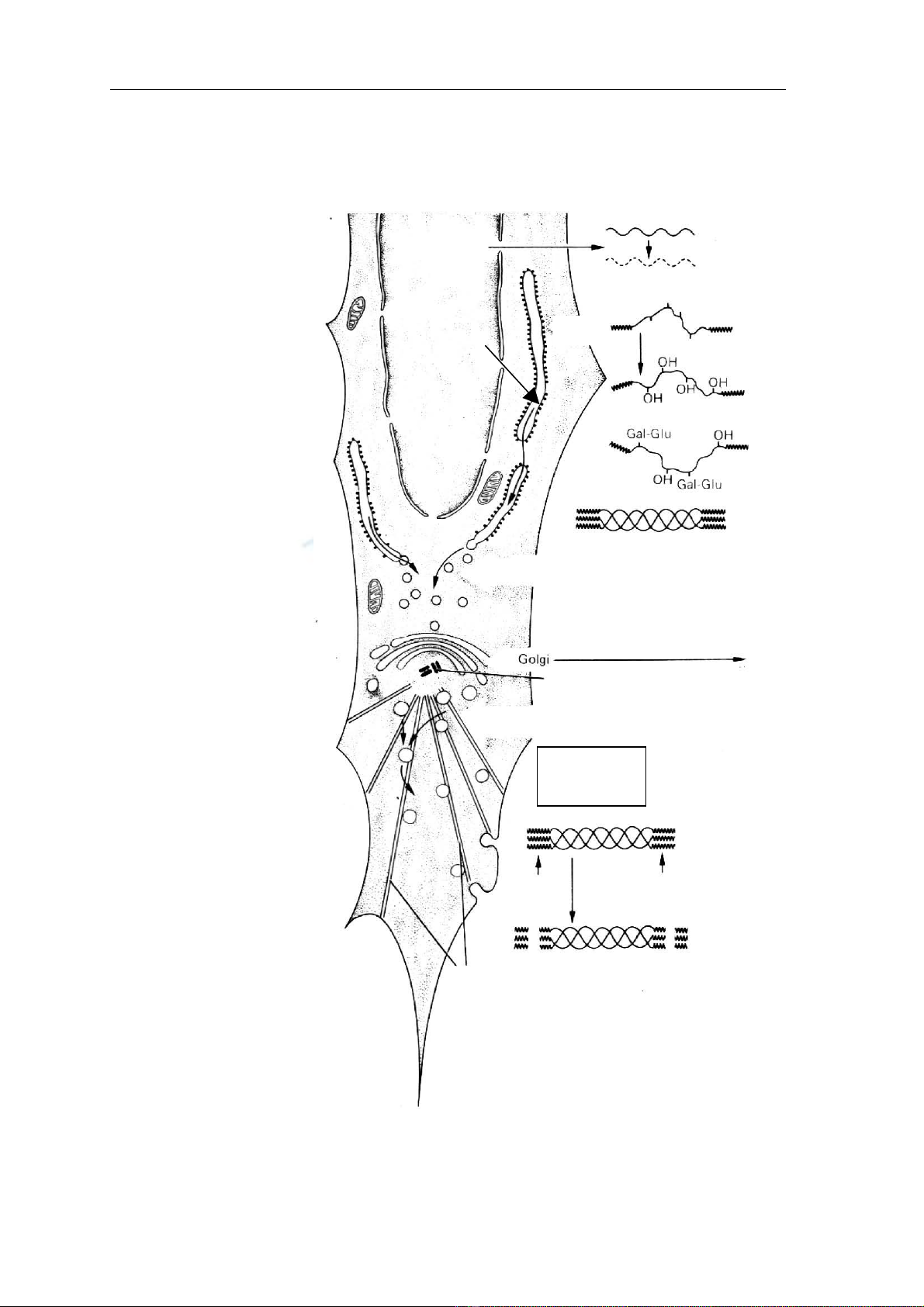
theo chuỗi tín hiệu ở đầu Hçnh 3: Så âäö täøng håüp såüi taûo keo tæì nguyãn baìo
såüi
11
Tropocollagen
Vi äúng
Càõt
Càõt
Ngoaûi baìo
Tiãön såüi
collagen
Procollagen
Såüi α
Nhán
Såüi RNA thäng tin cho mäùi
chuäùi α
Læåïi näüi
bao coï haût
Mä liãn kãút - Mä phäi
amino tận.
Ðây là một đoạn peptide có từ 20-50 acide amine giữ nhiệm vụ đưa toàn bộ chuỗi
polypeptide vào lưới nội bào. Trong một số trường hợp khác chúng có thể giữ nhiệm vụ đưa
chuỗi polypeptide ra ngoài màng tế bào, do đó còn có tên chuỗi tiết, chuỗi hướng dẫn (Hình
3).
Vậy tiền chuỗi ( gồm :chuỗi tín hiệu, đoạn peptide xa phía amino tận (20 acid amine),
chuỗi ( (1000-1050 acide amine) và đoạn peptide xa phía carboxyl tận (có khoảng 300 acide
amine).
Ở đoạn peptide xa, lysine là acide amine chiếm ưu thế, hydroxylysine có thể bị oxy
hoá khử amine carbon số 6 để tạo nên gốc aldehyde. Chính gốc aldehyde và hydroxylysine
làm cho các procollagene có thể liên kết với nhau một cách vững bền ở các ô lỗ lưới.
Mặt khác lysine cũng bị oxy hoá ở C5, ở vị trí này gốc OH của nó thường được gắn
với các đa đường phức tạp như N.acetyl glucose, N.acetyl glucosamine, N.acetyl
galactosamine. Các đa đường này là cơ sở để nối các procollagene cũng như đó là nơi đầu tiên
lắng đọng các muối hydroxyapatides trong quá trình cốt hoá chất căn bản xương. Ðoạn giữa
chính là chuỗi (, acide amine chiếm ưu thế trong chuỗi này là glycine, chiếm 1/3 tổng số các
acide amine, 2/3 còn lại là các acide amine khác và quan trọng nhất là proline và
hydroxyproline, sắp đặt có tính chất xen kẽ. Nếu gọi A và B là những acide amine khác với
glycine thì thứ tự của chuỗi có thể được minh hoạ bằng sơ đồ sau: A-gly-B-gly-A-gly-B-
Chính sự sắp xếp này cho phép đoạn giữa có cấu tạo helic bền vững. Hiện tại người ta
phát hiện ít nhất có 12 loại sợi (collagene khác nhau, về lý thuyết có thể suy đoán phải có
12x12x12=1728 type collagene khác nhau, tuy nhiên trong thực tế hiện nay người ta chỉ thấy
13 type collagene mà thôi, chúng được ký hiệu và phân bố ở các mô khác nhau.
Các procollagene sau khi được tổng hợp ở lưới nội bào sẽ được chuyển đến bộ Golgi,
tại đây các đoạn peptide xa về phía amine tận sẽ bị cắt. Phản ứng cắt cần enzime đặc hiệu, sau
đó chúng sẽ được chuyển vào các túi chứa procollagene và được chuyển đến sát màng tế bào,
các procollagene sẽ được đưa vào gian bào theo cơ chế ngược với cơ chế vi ẩm tế bào, ở gian
bào, một lần nữa các tiền sợi ( bị cắt về phía carboxyl tận và procollagene trở thành
tropocollagene.
12
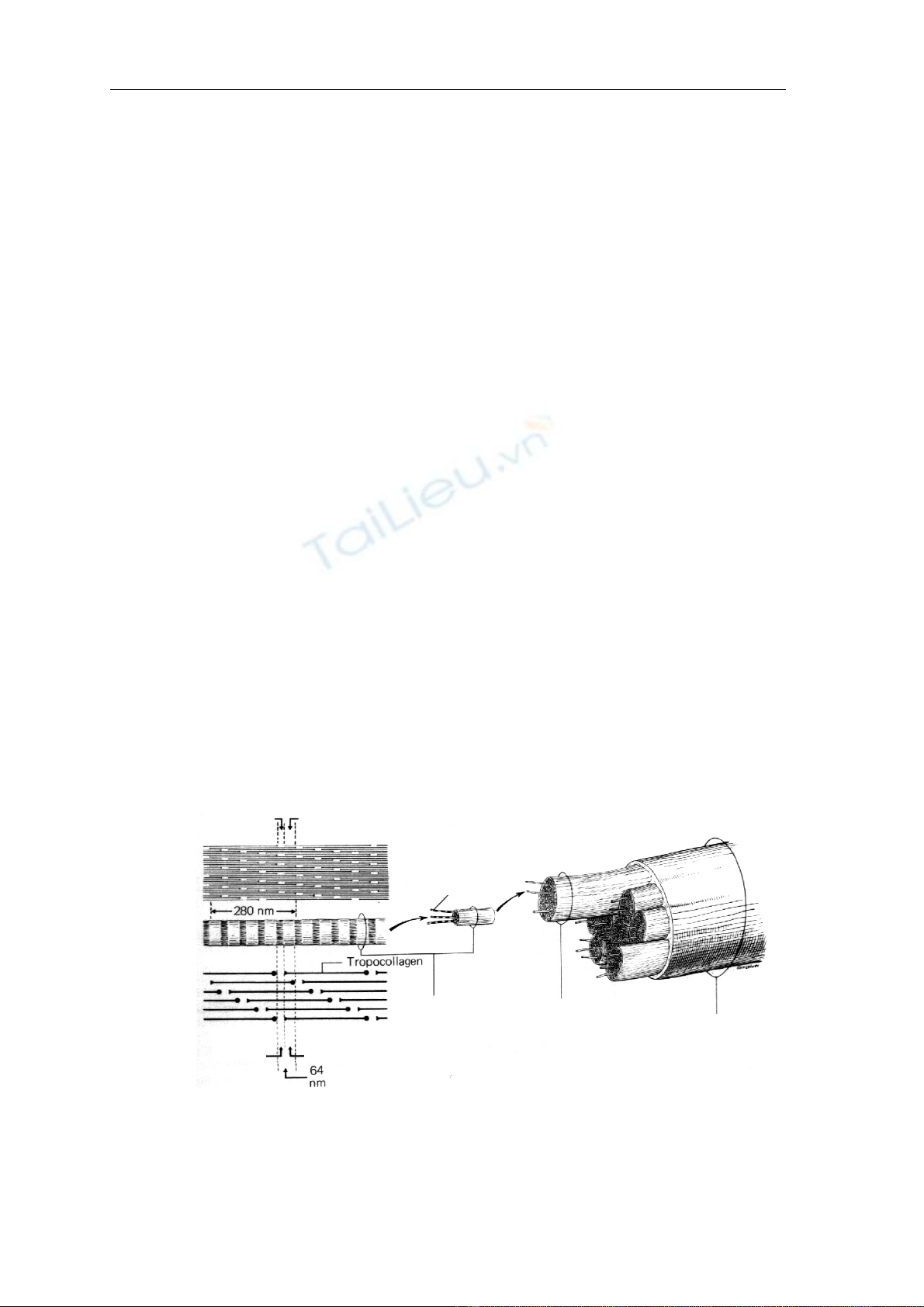
Boï såüi taûo keo
Såüi taûo keo
Vi såüi taûo keo
Bàng täúi
Bàng täúi
Chu kyì
Hçnh 4: Så âäö cáúu truïc boï såüi taûo keo,
såüi taûo keo, vi såüi taûo keo




![Câu hỏi ôn tập hóa sinh thực phẩm [chuẩn nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2018/20181019/ayunn123/135x160/2711539952639.jpg)
![Ôn tập thực hành môn Thực vật [chuẩn nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2015/20151028/ngochunghcm/135x160/1692190693.jpg)

![Định lượng Vitamin C: Bài 8 [Hướng Dẫn Chi Tiết]](https://cdn.tailieu.vn/images/document/thumbnail/2014/20140222/minhtriet16/135x160/7291393079428.jpg)





![Bài giảng Giáp xác chân mái chèo [mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20250927/lethihongthuy2402@gmail.com/135x160/92891759114976.jpg)



![Tài liệu học tập Chuyên đề tế bào [mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20250906/huutuan0/135x160/56151757299182.jpg)






