
SỰ PHÁT TRIỂN CHI CỦA ĐỘNG VẬT BỐN CHÂN
HÃY XEM XÉT CHÂN TAY CỦA BẠN. Nó có các ngón tay hoặc ngón chân ở một đầu, xương cánh
tay hoặc xương đùi ở đầu kia. Bạn sẽ không tìm thấy bất kỳ ai có ngón tay ở giữa cánh tay của họ. Cũng
nên xem xét sự khác biệt tinh tế nhưng rõ ràng giữa bàn tay và bàn chân của bạn. Nếu ngón tay của bạn
được thay thế bằng ngón chân, bạn chắc chắn sẽ biết điều đó. Bất chấp những khác biệt này, xương bàn
chân của bạn tương tự như xương bàn tay của bạn. Dễ dàng nhận thấy rằng chúng có chung một khuôn
mẫu. Và cuối cùng, hãy nhận thấy rằng cả hai bàn tay của bạn có kích thước tương tự nhau, cũng như cả
hai bàn chân của bạn. Những hiện tượng phổ biến này đặt ra những câu hỏi hấp dẫn đối với nhà sinh học
phát triển. Làm thế nào mà động vật có xương sống có bốn chi mà không phải sáu hoặc tám ? Làm thế
nào mà ngón út phát triển ở một cạnh của chi và ngón cái ở bên kia ? Làm thế nào để chi trước phát triển
khác với chi sau ? Làm thế nào kích thước chi có thể được điều chỉnh chính xác như vậy ? Có một bộ cơ
chế phát triển được bảo tồn nào có thể giải thích tại sao bàn tay của chúng ta có năm ngón, cánh gà có ba
ngón và móng ngựa một ngón không ?
Tổng quan
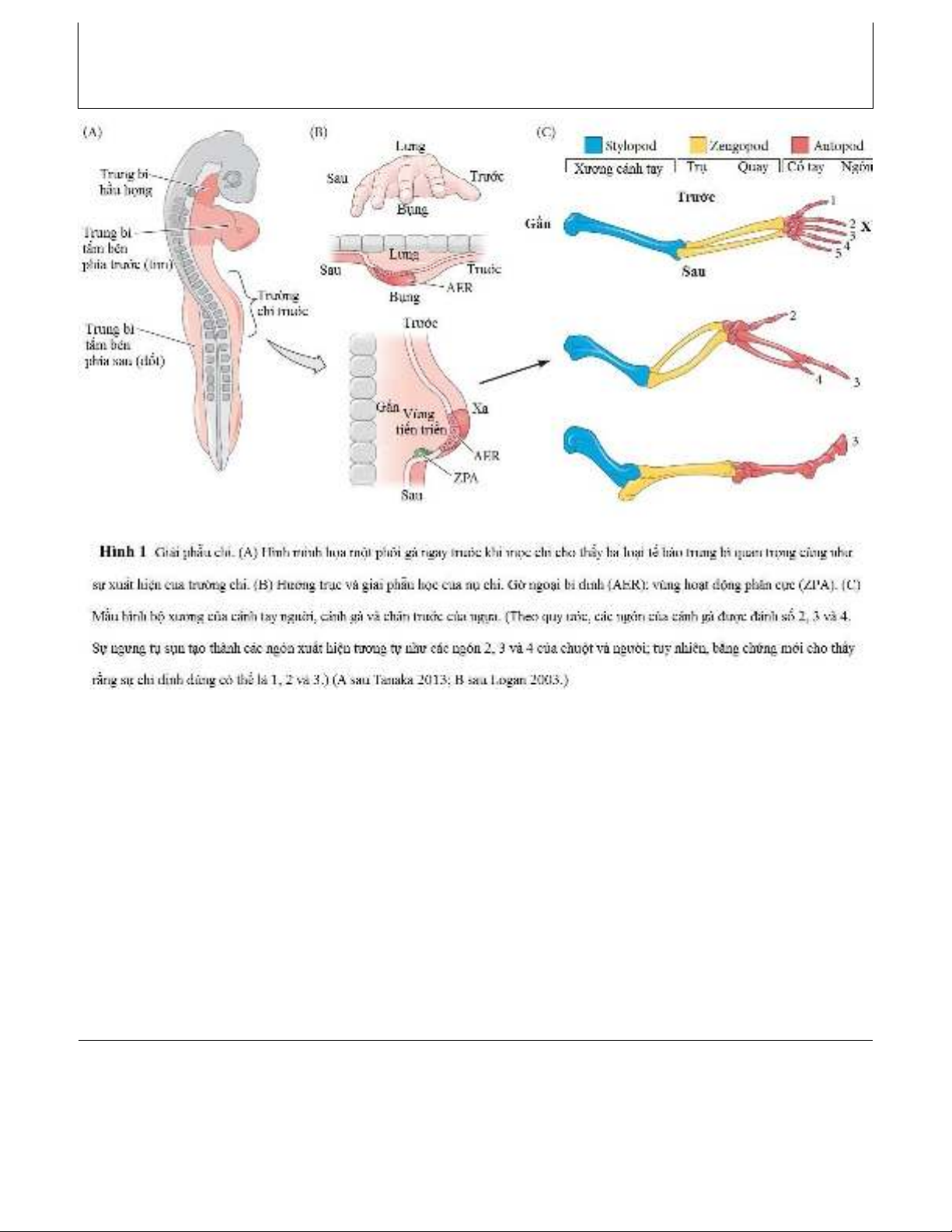
Được thiết kế để vận động trên cạn, tứ chi có khớp nối giữa các xương và một bộ ngón ở đầu xa của
chúng. Chi bắt đầu như là một “nụ” mô ở các mặt của phôi khi các tế bào từ trung bì khúc cơ và trung bì
tấm bên đang phát triển di chuyển đến và tăng sinh trong trường chi giả định, trung mô chi. “Vùng tiến
triển” có tính tăng sinh này được bao phủ bởi ngoại bì, với sự dày lên ở đầu xa được gọi là gờ ngoại bì
đỉnh. Tín hiệu Fgf8 từ gờ ngoại bì này đối kháng với axit retinoic có nguồn gốc từ cạnh sườn, bắt đầu một
vòng phản hồi dương tính với tín hiệu trung mô (Fgf10 và Wnt) để thúc đẩy nụ chi phát triển. Fgf8 chỉ
định trung mô phía sau thành “vùng hoạt động phân cực”, vùng này tiết ra Shh để thiết lập trục trước-sau
(ngón cái- út) của chi. Tín hiệu Wnt xác định trục lưng-bụng (mu bàn tay-lòng bàn tay). Quá trình tạo
xương được kiểm soát bởi một mô hình tự tổ chức “kiểu Turing” thông qua các tương tác của các chất tạo
hình thái. Ở một số loài động vật, mô màng sơ khai giữa các ngón vẫn còn được giữ lại; ở những loài
khác, nó chết thông qua quá trình apoptosis qua trung gian BMP. Mỗi hệ thống tín hiệu chi ảnh hưởng
đến sự biểu hiện khác biệt của các gene Hox dọc theo mỗi trục, việc sửa đổi chúng hỗ trợ sự phát triển của
vây thành các ngón tay.
Giải phẫu chi
Như tên gọi, động vật có xương sống bốn chi (lưỡng cư, bò sát, chim và động vật có vú). Xương của bất
kỳ chi của động vật bốn chân nào — có thể là cánh tay hoặc chân, cánh hoặc chân chèo — xét đến một
styopod ở gần (xương cánh tay / xương đùi) liền kề với thành cơ thể, một zeugopod (trụ-quay / chày-mác)
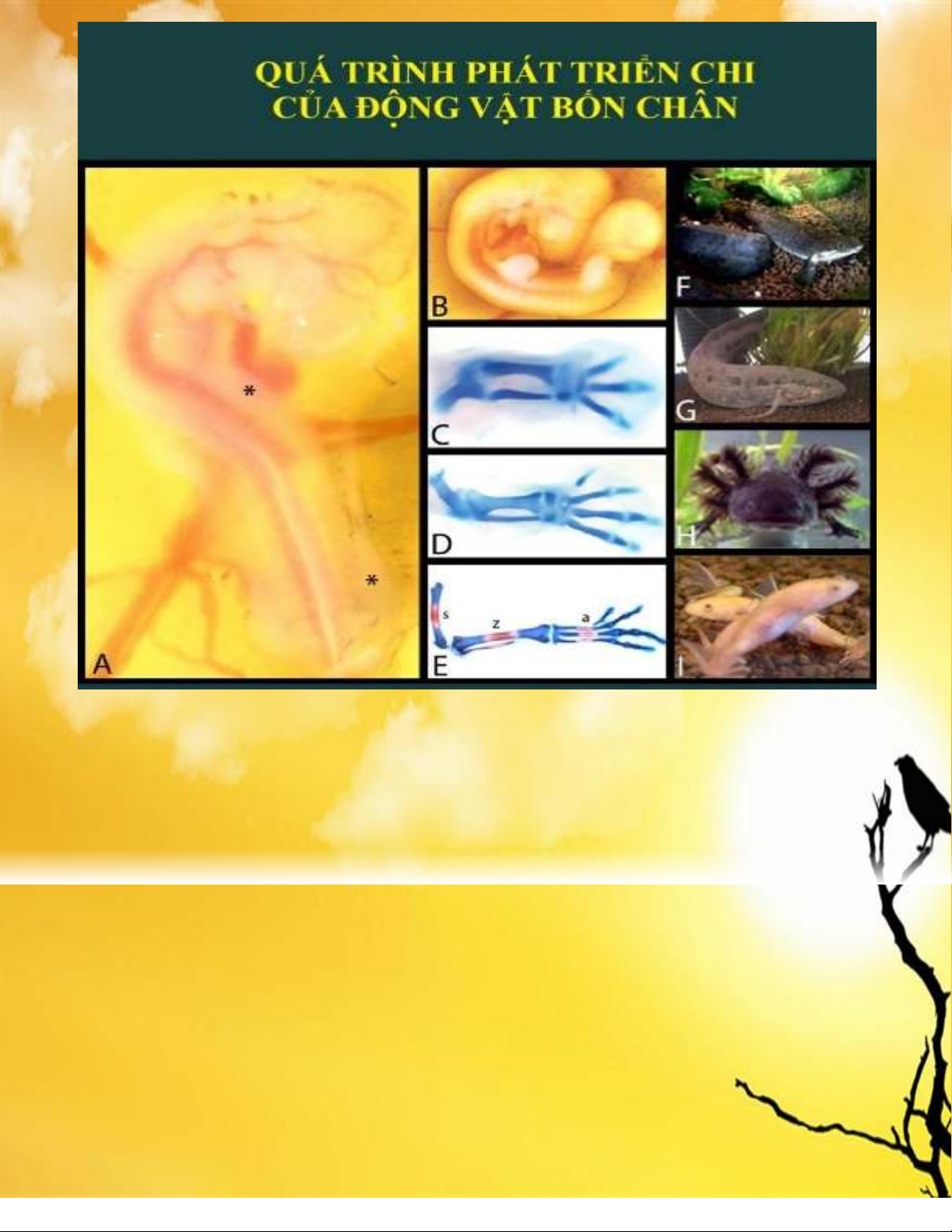
ở vùng giữa và một autopod ở xa (cổ tay-ngón tay / cổ chân-ngón chân) 1(HÌNH 1). Ngón tay và ngón
chân có thể được gọi là các đốt ngón (phalange) hoặc nói chung hơn là ngón (digit). Thông tin vị trí cần
thiết để tạo ra một chi phải hoạt động trong một hệ tọa độ ba chiều 2:
1Những thuật ngữ này có thể khó nhớ, nhưng biết nguồn gốc từ của chúng có thể hữu ích. Stylo = như một cây cột;
zeugo = khớp nối; auto = bản thân; pod = chân
2Thực tế, nó là một hệ bốn chiều, trong đó thời gian là trục thứ tư. Các nhà sinh học phát triển đã quen với việc nhìn
thiên nhiên trong bốn chiều.

• Chiều đầu tiên là trục gần-xa (“gần-xa”; nghĩa là từ vai tới ngón tay hoặc từ hông tới ngón chân). Các
xương của chi được hình thành bởi quá trình hóa hạch nội sụn. Ban đầu chúng là sụn, nhưng cuối cùng
hầu hết sụn được thay thế bằng xương. Bằng cách nào đó, các tế bào chi phát triển khác nhau ở giai đoạn
đầu của quá trình phát sinh hình thái chi (khi chúng tạo ra stylopod) so với ở các giai đoạn sau (khi chúng
tạo ra autopod).
• Chiều thứ hai là trục trước - sau (ngón cái đến ngón út). Ngón tay út hoặc ngón chân út của chúng ta
đánh dấu đầu phía sau và ngón cái hoặc ngón chân cái của chúng ta ở đầu phía trước. Ở con người, rõ
ràng là mỗi bàn tay phát triển như một hình ảnh phản chiếu của tay kia. Người ta có thể tưởng tượng
những cách sắp xếp khác — chẳng hạn như ngón cái phát triển ở bên trái của cả hai tay — nhưng những
kiểu này không xảy ra.
• Cuối cùng, các chi có trục lưng-bụng: lòng bàn tay (bụng) của chúng ta dễ dàng phân biệt với mu bàn
tay (lưng).
Nụ chi
Dấu hiệu đầu tiên có thể nhìn thấy được của sự phát triển chi là sự hình thành các khối phồng ở hai bên
gọi là các nụ chi ở các vị trí giả định của chi trước và chi sau (Hình 2A). Nghiên cứu lập bản đồ số phận
trên sa giông, do phòng thí nghiệm của Ross Granville Harrison tiên phong (xem Harrison 1918, 1969),
cho thấy trung tâm của đĩa tế bào này trong vùng thân thể của trung bì tấm bên thường tự phát sinh chi.
Liền kề với nó là các tế bào sẽ hình thành mô cạnh sườn quanh phế quản (xung quanh chi) và xương đòn
vai. Tuy nhiên, nếu tất cả các tế bào này được tách ra khỏi phôi, thì một chi vẫn sẽ hình thành (mặc dù hơi
muộn hơn) từ một vòng tế bào bổ sung bao quanh khu vực này nhưng thông thường sẽ không hình thành
chi. Nếu vòng tế bào xung quanh này được bao gồm trong mô bị cắt bỏ, thì sẽ không có chi nào phát triển.
Vùng lớn hơn này, đại diện cho tất cả các tế bào trong vùng có khả năng tự hình thành một chi, là trường
chi.
Các tế bào tạo nên nụ chi có nguồn gốc từ trung bì tấm bên ở phía sau, kế cận các đốt và ngoại bì bên trên
của nụ chi. Các tế bào trung mô tấm bên di chuyển bên trong các trường chi để hình thành các tế bào tiền
thân của bộ xương chi, trong khi các tế bào trung mô từ các đốt ở cùng mức di chuyển đến để thành lập
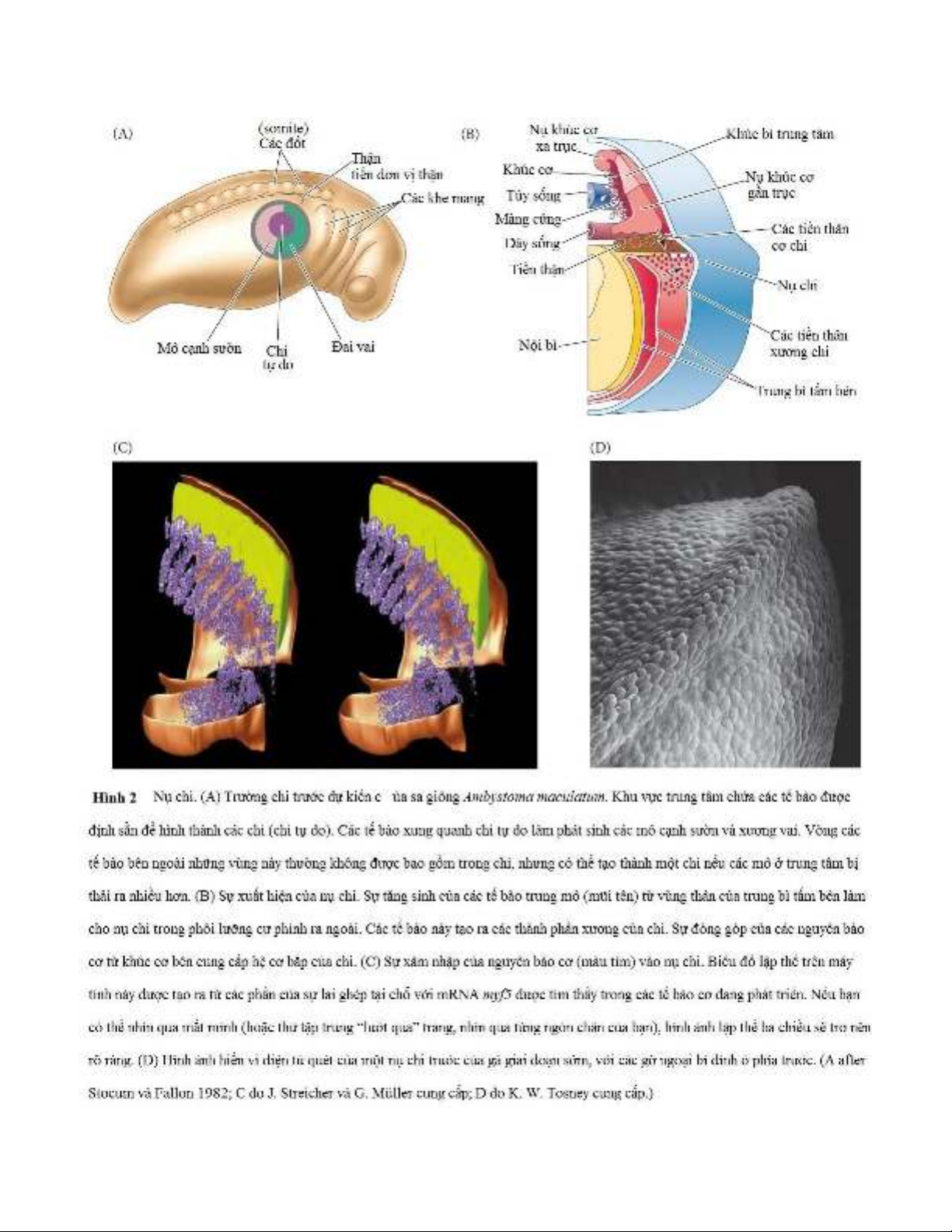
các tế bào tiền thân của cơ chi (Hình 2B, C). Quần thể không đồng nhất đang tích lũy này của các tế bào
trung mô tăng sinh bên dưới mô ngoại bì, tạo ra nụ chi.
Ngay cả nụ chi sớm cũng sở hữu cách thức tổ chức riêng của nó sao cho hướng phát triển chính xảy ra
dọc theo trục từ gần đến xa (từ các đốt somite đến ngoại bì), với sự phát triển ít hơn xảy ra dọc theo trục





![Giáo trình trang bị công nghệ và cấp phôi tự động C10 [chuẩn nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2011/20110727/trantrungthai/135x160/chuong_10_6319.jpg)








![Hướng dẫn giải chi tiết bài tập phân li, phân li độc lập: Tài liệu [mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20251204/lethu2868@gmail.com/135x160/84711764814448.jpg)



![Bài tập Đa dạng thế giới sống [kèm đáp án/ hướng dẫn giải]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20251123/thaohoang9203@gmail.com/135x160/5861763951302.jpg)




