ĐỘT BIẾN GEN
GV: Nguyễn Thị Ngọc Yến
Trang web slide bài giảng bộ môn Vi – ký sinh: https://sites.google.com/site/bomonvikysinh/
Mục tiêu
Đột biến là gì? Phân loại đột biến Nguyên nhân và hậu quả Các yếu tố gây đột biến Bệnh do đột biến ở người Phương pháp chọn lọc đột biến
Khái niệm
thư, lão hóa
o Đột biến dòng mầm (xảy ra trong giao tử): có tính di
truyền, là gen lặn, cần 2 bản allen để biểu hiện
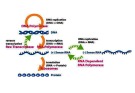
Biến đổi di truyền (thay đổi trình tự nucleotid/ gen) xảy ra đột ngột tạo ra protein đột biến ảnh hưởng sự tổng hợp sản phẩm gen o Đột biến sinh dưỡng: không di truyền, có thể gây ung
Tần số đột biến
chép, 1 lần phân bào, 1 giao tử, 1 tế bào
Tần số đột biến được đánh giá căn cứ trên 1 lần sao
Mỗi gen có tần suất ổn định, sự khác nhau về tỉ lệ đột
Tần số đb từng gen thấp nhưng tổng đb ở người là con
biến là do: o Kích thước gen: lớn dễ đột biến o Điểm nóng: gen nằm trên vùng NST nhạy cảm
số lớn, ý nghĩa trong tiến hóa: o Gen: 10-6/gen/thế hệ o Giao tử: 10-6.5.104 gen = 1/20 o Hợp tử: 1/20.2 = 1/10
Đột biến: lợi và hại
Vùng mã hóa: thay đổi đặc tính protein Vùng không mã hóa: thay đổi trình tự nu không gây
khác biệt tính trạng đb tích tụ trong gen
Kỹ thuật cảm ứng đột biến Đột biến điểm định hướng
Phân loại
Thêm 1 cặp nu
ĐB lệch khung
Mất 1 cặp nu
ĐB điểm
Chuyển vị
Thay thế cặp nu
Đột biến
Đảo chuyển
ĐB đa điểm
Đột biến điểm
ĐB thay thế cặp nucleotid
Nguyên nhân
Hậu quả
Trong vùng mã hóa
PurinPurin
o Đb lặn (cùng nghĩa): mã hóa cùng aa không ảnh hưởng
Chuyển vị
Pyrimidinpyrimidin
o Đb lệch nghĩa: mã hóa aa khác ảnh hưởng ít/ nhiều
Purinpyrimidin
o Đb vô nghĩa: mã hóa codon kết thúc dừng sớm/ cụt
Đảo chuyển
Pyrimidinpurin
Trong vùng không mã hóa: ảnh hưởng phiên mã, dịch mã, mức xoắn bện của ADN
Đb lùi (hồi biến)
ĐB thay thế cặp nucleotid
ĐB lặn
ĐB vô nghĩa
ĐB lệch nghĩa
Đột biến lệch nghĩa
ĐB lệch khung
Nguyên nhân
Hậu quả
Xảy ra/ vùng mã hóa
Thay đổi khung đọc từ vị trí đb về sau
Chèn 1 nu
Chứa stop codon ngừng sớm
Mất 1 nu
Chèn + mất/ ADN
Thêm, bớt đồng thời sửa khung đọc giữa 2 điểm đb
Bình thường
Mất nu
Thêm nu
Thay thế nu
ĐB đa điểm
Nguyên nhân
Hậu quả
Xảy ra/ vùng mã hóa
Sao chép các yếu tố lặp lại
Lệch khungthay đổi pro
Transposon
Ngưng phiên mã, dịch mã
Gen nhảy
Retrotransposon
ADNARNADN
Tác động đến hơn 1 base gây xóa/ chèn Không hồi biến (ổn định)
Nguyên nhân đột biến
- ĐB tự nhiên - ĐB cảm ứng (nhân tạo)
Đb tự nhiên
Nguyên nhân ĐB tự nhiên
xác định nguồn gốc, tỉ lệ đb thấp
Xảy ra ngẫu nhiên với tần số nhất định 10-10, không
Nguyên nhân: gắn sai base/sao chép đb điểm Khắc phục: ADN pol hoạt tính exonuclease 3’5’ Phân loại o Hỗ biến o Khử amin o Đb lệch khung o Oxy hóa phá hủy do các gốc oxy
ĐB tự nhiên: Hỗ biến
o Keto (C=O) enol (C-OH): T bắt cặp G và ngược lại o Amino (NH2) imino (=NH): C bắt cặp A và ngược lại
Các base có khả năng hoán chuyển giữa 2 dạng:
chuyển vị G-C A-T
ĐB tự nhiên: Khử amin
o C U: chỉnh lại bằng loại U (vì ADN không có U) o 5-methylcytosin T: không chỉnh được. C tại các
trình tự CpG dễ bị methyl hóa “điểm nóng” đb
Phản ứng thủy phân
ĐB tự nhiên: Lệch khung
Chèn/ mất trên 1 sợi Nguyên nhân: lỗi của ADN pol khi sao chép các đoạn lặp lại của nucleotid. Tính chính xác của pol do hoạt tính exonuclease 3’5’
ĐB lệch khung: Oxy hóa
Gốc oxy trong tế bào (tia phóng xạ) bắt cặp sai với A, tạo đảo chuyển G-C A-T
ĐB cảm ứng (nhân tạo)
ĐB cảm ứng
Tác nhân làm tăng tần số đb cao hơn tự nhiên là tác nhân gây đb: phóng xạ, tia X, tử ngoại…
Tác nhân hóa học Tác nhân bức xạ
ĐB cảm ứng: Tác nhân hóa học
Base đồng đẳng Tác nhân thay đổi đặc tính bắt cặp: chất khử amin và alkyl hóa Tác nhân chèn vào ADN Tác nhân thay đổi cấu trúc ADN
Base đồng đẳng
Cấu trúc ~ purin/pyrimidin gắn vào ADN gây chuyển vị hay hỗ biến tự phát
o Dạng keto (BU) = T: bắt cặp A o Dạng enol (BU*) = C: bắt cặp G Chuyển vị A-T thành G-C và ngược lại
Aminopurin (AP) = A: bắt cặp T chuyển vị A-T
Bromouracil (BU):
thành G-C
Bromouracil
Chuyển vị A-TG-C
Keto (BU)=T
Enol (BU*)=C
*
*
Aminopurin
Chuyển vị A-TG-C
Chất khử amin
Base ngoại: Khởi động cơ chế sửa chữa
Tác nhân: acid nitrơ và hydroxylamin
Chất khử amin
là “điểm nóng” cho đb xảy ra (CpG site)
Metylcytosin ảnh hưởng đến hoạt tính phiên mã, cũng
Thay đổi cấu trúc, đặc tính bắt cặp
Chất alkyl hóa: methyl/ethyl hóa base, nhất là guanin hay các nhân pyrimidin dễ bị alkyl hóa o Nitrosoguanidin (NTG) o Methyl methan sulfonat (MMS) o Ethyl methan sulfonat (EMS)
Thay đổi cấu trúc, đặc tính bắt cặp
sai với thymin
Gắn (-CH3), (-C2H5) vào guanin = adenin bắt cặp
Liên kết chéo giữa các mạch của 1 hoặc các ADN làm
mất nucleotid
Thay đổi cấu trúc, đặc tính bắt cặp
khi sao chép làm đứt mạch
Mất purin do guanin bị alkyl hóa tạo lỗ hổng trên ADN,
Tác nhân chèn vào ADN
Gồm: proflavin, cam acridin, ethidium bromide Phân tử đa vòng phẳng tương tác và chèn vào ADN
Giãn sợi đôi ADN,
ADN polymerase
bị
“đánh lừa”, chèn base
vào chỗ trống,
làm
lệch khung ADN
Tác nhân thay đổi cấu trúc ADN
N-acetoxy-2-acetylaminoflourene (NAAAF)
Phân tử lớn, gây liên kết chéo trong và giữa các sợi:
Chất hóa học gây đứt sợi ADN: peroxid Hydrocarbon đa vòng: benzopyren
Bức xạ điện từ
Bức xạ ion hóa
Tia UV
ĐB cảm ứng: Tác nhân bức xạ
- Không có ngưỡng tác dụng - Tỉ lệ thuận liều lượng phóng xạ, không phụ thuộc cường độ và thời gian chiếu xạ
Bức xạ
Bức xạ ion hóa
Tia cực tím – BX không ion hóa
Phân loại
Tia X, tia gama, hạt phóng xạ α, β
UV-C (180-290nm): gây chết UV-B (290-320): gây đb chính UV-A (320): cận UV, ít gây đb
Cơ chế gây đb
Ion hóa các phân tử trên đường đi tạo gốc tự do tương tác với ADN, protein, tế bào (máu, tủy xương, niêm mạc)
Khả năng xuyên thấu thấp: tác động lên đơn bào và giao tử ADN hấp thu cực đại 257nm bước sóng tăng tần số đb
Tác dụng di truyền
Tạo thymin dimer (UV-B, UV-C) Ngăn phiên mã, sao chép, đb NST, gây chết nếu không sửa chữa Khắc phục: quang phục hồi
-Đứt 1 hoặc 2 sợi -Phá hủy các base -Liên kết chéo trong ADN hoặc ADN với protein -Hỏng bào quan, ngăn phân chia, làm chết tb
BX ion hóa
Tia UV
Cơ chế chống lại đột biến
•Đảo nghịch sai hỏng
•Loại bỏ sai hỏng
•Dung nạp sai hỏng
Đảo nghịch sai hỏng
Enzym phục hồi cấu trúc bình thường ko gãy khung Quang phục hồi: photolyase xúc tác quá trình cắt loại các pyrimidin dimer. Không có ở động vật.
Đảo nghịch sai hỏng
O6-methylguanin, O4-methylthymin,
Làm mất nhóm alkyl nhờ methyltransferase bằng cách chuyển gốc methyl từ oxy trên ADN sang cystein trên enzym. PƯ ko thuận nghịch, enzym mất hoạt tính. Lỗi O6- gồm ethylguanin…
Nối các chỗ đứt do tia X và peroxid bằng ligase
Loại bỏ sai hỏng
Cắt base (N-glycosylase) Cắt nucleotid (NER)
Loại bỏ sai hỏng
Sửa chữa lệch đôi: Xảy ra sau khi sao chép để “đáh vần kiểm tra” cuối
cùng. Protein quét ADN tìm base bắt cặp sai
Tăng độ chính xác lên 100-1000 lần
Dung nạp sai hỏng
Không sửa chữa, sao chép sai hỏng và tiếp tục sống Sửa chữa bằng tái tổ hợp Sửa chữa bằng đột biến: “đột biến hoặc là chết”
Sửa chữa bằng
tái tổ hợp: lấp
khuyết sợi con
không dimer, cơ
chế chính xác
Tính trạng và protein đột biến
Bệnh do đột biến
di truyền được
SV đột biến: SV biểu hiện những tính trạng bất thường
Rối loạn chuyển hóa bẩm sinh (bệnh phân tử): nguồn gốc từ những sai sót mức độ phân tử: “một gen một enzym”
Bệnh do đột biến
Thiếu máu hồng cầu hình liềm Thiếu máu Địa Trung Hải Thiếu máu hồng cầu hình bia Bệnh máu không đông Teo cơ Dunchenne Không dung nạp lactose Phenylketon niệu Porphyria
Đột biến gen và Ung thư
Ung thư
Nguyên nhân
Bất hoạt gen kìm hãm
Tăng hoạt gen kích thích
Proto-oncogen Oncogen
ĐB trội
ĐB lặn
Sự tăng sinh tế bào không kiểm soát
Ung thư
Gen ung thư do retrovirus tải nạp được xen vào cạnh gen tiền ung thư tb chủ
Ung thư
Biến đổi gen tiền ung thư thành ung thư Đb điểm hoặc mất đoạn Khuếch đại gen Chuyển đoạn NST
Đột biến ở gen ức chế khối u
Gen Rb
tạo ức retinoplasma chế sự phát triển ung thư mắt p53 Gen
tạo p53 ức protein chế nhiều dạng ung thư
Virus gây UT bằng cách cô lập sp các gen này
Hệ thống chọn lọc đột biến
•PP đề kháng •PP làm giàu chậm •PP làm giàu hạn chế •PP làm giàu nhờ penicillin •PP lọc •PP in








![Tài liệu giảng dạy Sinh học và di truyền [mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2026/20260323/hoatudang2026/135x160/42181774414220.jpg)

![Giáo trình Công nghệ vi sinh (Nghề Công nghệ sinh học TC/CĐ) - Trường Cao đẳng Đà Lạt [Mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2026/20260224/hoacattuong2026/135x160/87621772161812.jpg)






![Giáo trình Vi sinh vật học môi trường Phần 1: [Thêm thông tin chi tiết nếu có để tối ưu SEO]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20251015/khanhchi0906/135x160/45461768548101.jpg)





