HIV-1
(no Vpx)
SIVsm/HIV-2
(with Vpx)
Cytoplasm
Reverse
transcription
Human myeloid-
lineage cell
Permissive
infection
Restrictive
infection
Vpx
Vpx
Reverse
transcription
SAMHD1
Proteasome
Ubiquitin
DCAF1
CUL4A/
DDB1
Vpx
E3
D
V
SAMHD1
i
t
ti
n
ti
Degradation
SAMHD1: a new insight into HIV-1 restriction in
myeloid cells
St Gelais and Wu
St Gelais and Wu Retrovirology 2011, 8:55
http://www.retrovirology.com/content/8/1/55 (8 July 2011)

VIEWPOIN T S Open Access
SAMHD1: a new insight into HIV-1 restriction in
myeloid cells
Corine St Gelais and Li Wu
*
Abstract
Human myeloid-lineage cells are refractory to HIV-1 infection. The Vpx proteins from HIV-2 and sooty mangabey
SIV render these cells permissive to HIV-1 infection through proteasomal degradation of a putative restriction
factor. Two recent studies discovered the cellular protein SAMHD1 to be this restriction factor, demonstrating that
Vpx induces proteasomal degradation of SAMHD1 and enhances HIV-1 infection in myeloid-lineage cells. SAMHD1
functions as a myeloid-cell-specific HIV-1 restriction factor by inhibiting viral DNA synthesis. Here we discuss the
implications of these findings in delineating the mechanisms of HIV-1 restriction in myeloid-lineage cells and the
potential role of Vpx in lentiviral pathogenesis.
Introduction
Myeloid-lineage cells, including monocytes, dendritic
cells (DCs) and macrophages, play a multifaceted role in
HIV-1 initial infection and viral dissemination; however,
these cell types are restrictive to post-entry HIV-1 infec-
tion in vitro [1,2]. For gene therapy purposes, transduc-
tion of human DCs with an HIV-1-derived lentiviral
vector can be significantly enhanced by preincubation
with virus-like particles derived from SIV [3]. Subse-
quent studies indicated that Vpx proteins from sooty
mangabey SIV (SIVsm) and HIV-2 lineages efficiently
enhanceHIV-1infectioninhumanDCsandpromote
the accumulation of full-length viral DNA [4]. Further
studies from several laboratories suggested that Vpx,
similar to HIV-1 Vpr, interacts with the DCAF1 compo-
nent of the CUL4A/DDB1 and E3 ubiquitin ligase com-
plex (reviewed in [5,6]). However, only SIVsm/HIV-2
Vpx can efficiently enhance HIV-1 infection in DCs and
macrophages [5]. These studies led to the hypothesis
that Vpx targets a putative HIV-1 restriction factor for
proteasomal degradation in myeloid cells through the
E3 ubiquitin ligase complex [5,6], prompting the search
for the unknown HIV-1 restriction factor in human
myeloid cells that is counteracted by Vpx.
SIVsm, SIVsm-derived rhesus macaque SIV (SIVmac),
and HIV-2 encode both Vpr, a homologue of the HIV-1
Vpr protein, and Vpx, a protein unique to the SIVsm
lineage. vpx has likely evolved via duplication of the pri-
mate lentivirus vpr gene [5]. Early studies have demon-
strated that macaques infected with Vpx-defective
SIVmac or SIVsm had decreased viremia, impaired viral
replication, and slower AIDS progression compared to
wild-type SIV-infected animals, thus revealing the
importance of Vpx in SIV pathogenesis [7,8]. The
important role of Vpx in lentiviral infection in myeloid-
lineage cells in vitro and in vivo indicates that Vpx is
not merely a functional copy of Vpr, but may possess a
unique function. Although Vpx has been reported to
facilitate nuclear import of viral DNA [5], the precise
function of Vpx in lentiviral pathogenesis remains to be
defined.
New findings and discussion
Using mass spectrometry, Laguette et al. identified
SAMHD1 as a novel Vpx-interacting protein purified
from differentiated human monocytic THP-1 cells that
express tagged Vpx [9]. The rationale for using THP-1
cells was based on the previous work that differentiated
THP-1 cells can be rendered more permissive to HIV-1
infection by transduction of SIVsm/HIV-2 Vpx-contain-
ing virus-like particles derived from SIVmac [5].
SAMHD1 is expressed in non-permissive cells, including
THP-1 cells, primary monocytes, monocyte-derived
macrophages and DCs, while permissive CD4
+
T cells
and monocytic U937 cells do not express endogenous
SAMHD1 [9], suggesting an inverse correlation between
SAMHD1 expression and permissiveness to HIV-1
* Correspondence: wu.840@osu.edu
Center for Retrovirus Research, Department of Veterinary Bioscience, The
Ohio State University, 1900 Coffey Road, Columbus, OH 43210, USA
St Gelais and Wu Retrovirology 2011, 8:55
http://www.retrovirology.com/content/8/1/55
© 2011 St Gelais and Wu; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative
Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and
reproduction in any medium, provided the original work is properly cited.
infection. Moreover, silencing of SAMHD1 in non-
permissive cells (THP-1 cells and DCs) alleviates HIV-1
restriction, and over-expression of SAMHD1 in permis-
sive cells (HeLa cells and U937 cells) inhibits HIV-1
infection [9].
By contrast, Hrecka and colleagues identified
SAMHD1 from HEK 293T cells expressing tagged Vpx
in a proteomic screen using multidimensional protein
identification technology [10]. They demonstrated that
Vpx relieves the inhibition of HIV-1 infection in mono-
cyte-derived macrophages by mediating proteasome-
dependent degradation of SAMHD1 through the
CUL4A/DCAF1 E3 ubiquitin ligase [10]. Both studies
confirmed that Vpx interacts with SAMHD1 and
induces proteasomal degradation of SAMHD1 in THP-1
cells or macrophages, which can be restored by treat-
ment with a proteasome inhibitor [9,10].
The HD domains have putative nucelotidase and
phosphodiesterase activities, and the highly conserved
histidine (H) and aspartic acid (D) residues are critical
for catalytic activity [11]. Indeed, Laguette et al. showed
that over-expression of a HD domain mutant SAMHD1
in U937 cells fails to restrict HIV-1, suggesting that the
phosphodiesterase activity of the HD domain is impor-
tant for the restriction function of SAMHD1. Further
analysis revealed that SAMHD1 blocks HIV-1 reverse
transcription, as silencing SAMHD1 in THP-1 cells [9]
and macrophages [10] increases the levels of viral DNA.
Together, these studies suggested that SAMHD1 is the
myeloid-cell specific HIV-1 restriction factor counter-
acted by Vpx [9,10] (Figure 1).
The new findings by Laguette et al. and Hrecka et al.
have opened the door towards understanding the poten-
tial role of SAMHD1 in lentiviral pathogenesis.
SAMHD1-mediated HIV-1 restriction in myeloid-lineage
cells protects these cells from efficient HIV-1 infection,
which likely prevents an innate immune response trig-
geredbyHIV-1.Bycontrast,SIVsmandHIV-2encode
HIV-1
(no Vpx)
SIVsm/HIV-2
(with Vpx)
Cytoplasm
Reverse
transcription
Human myeloid-
lineage cell
Permissive
infection
Restrictive
infection
Vpx
Vpx
Reverse
transcription
SAMHD1
Proteasome
Ubiquitin
DCAF1
CUL4A/
DDB1
Vpx
E3
D
V
SAMHD1
it
ti
n
ti
Degradation
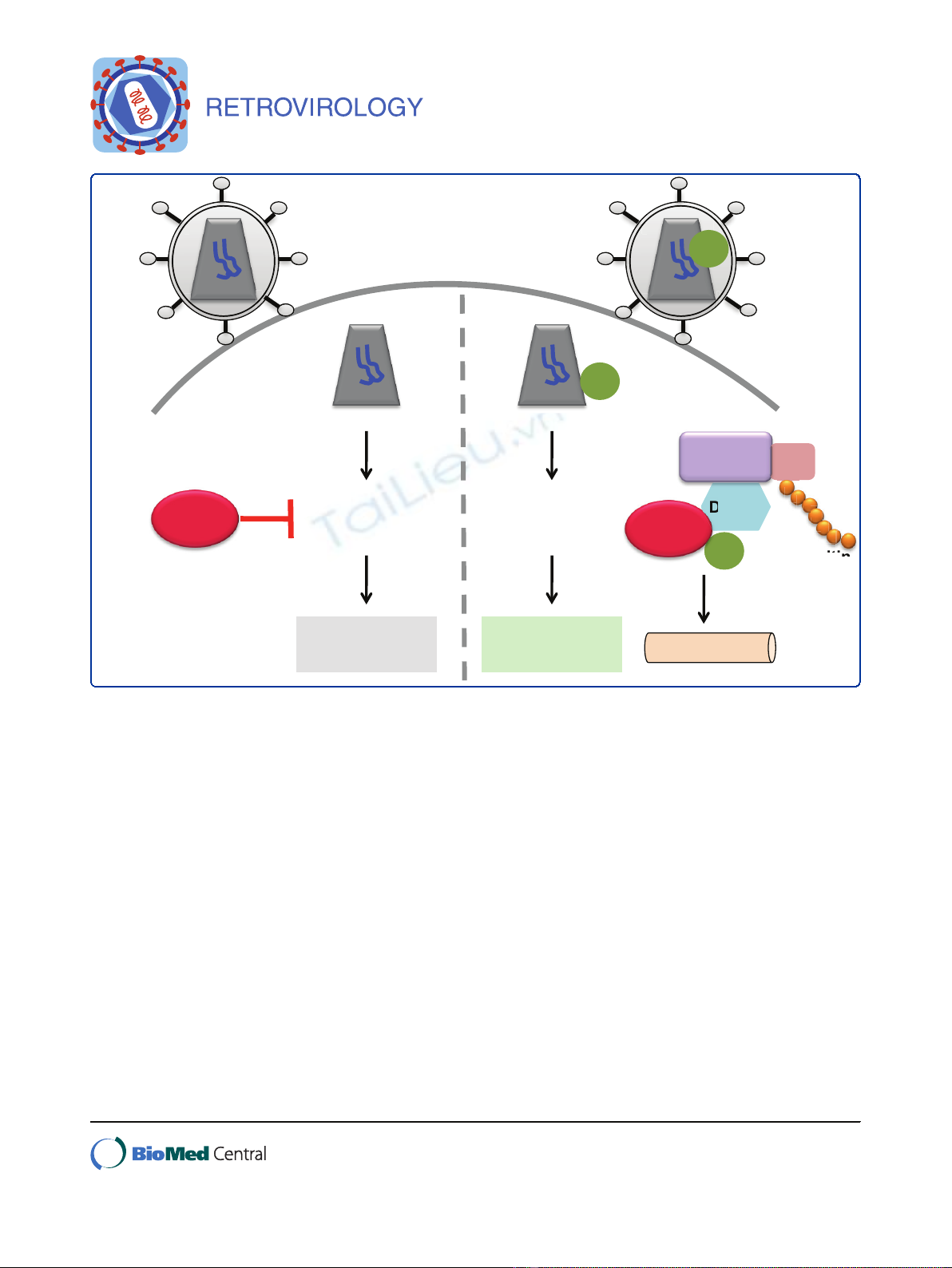
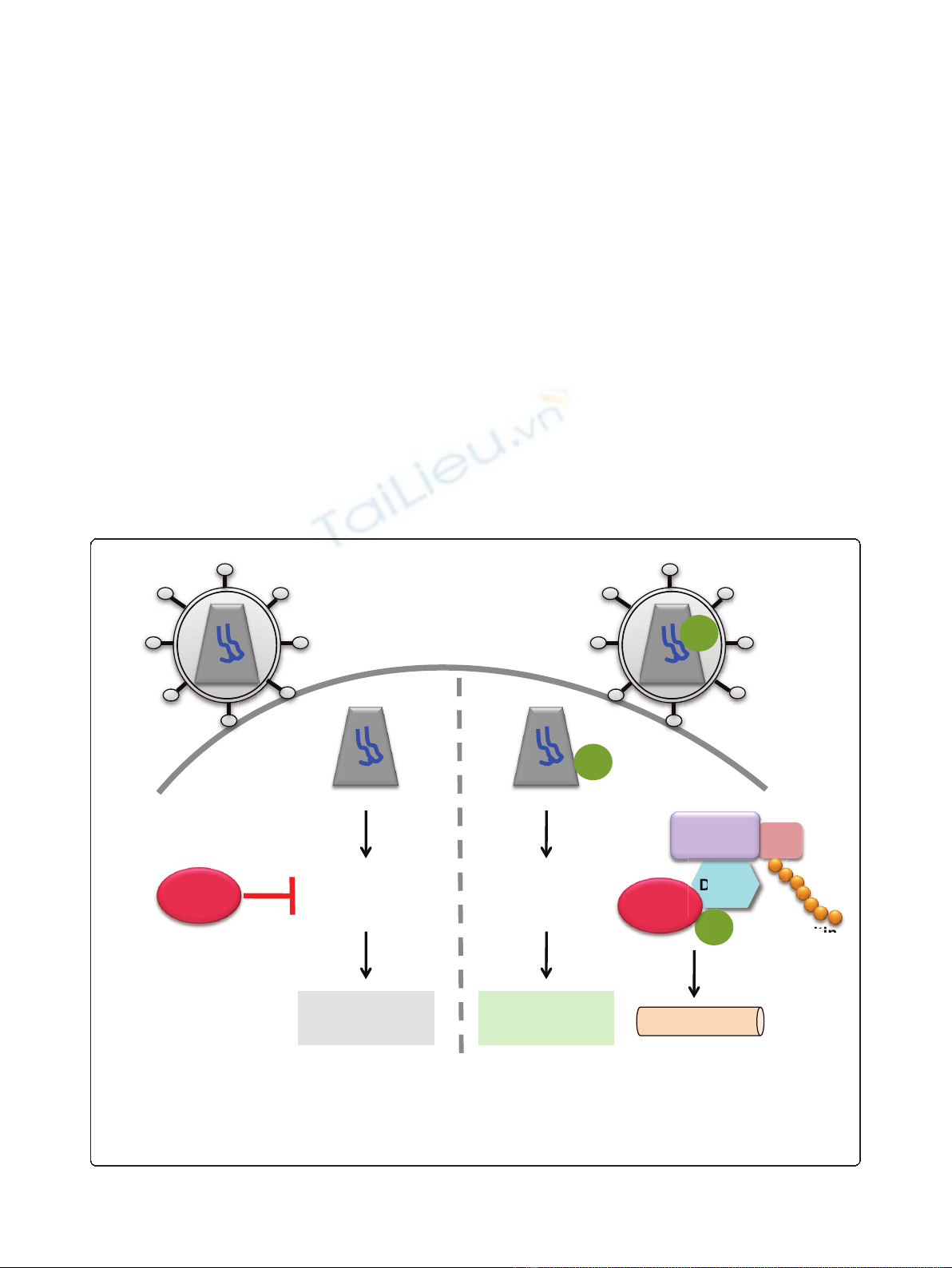
Figure 1 Vpx interacts with the E3 ubiquitin ligase complex to target the restriction factor SAMHD1 for proteasomal degradation.
Human myeloid-lineage cells that are non-permissive to HIV-1 infection express high levels of SAMHD1, which appears to act early in infection
at the reverse transcription step. HIV-1 has not evolved a viral antagonist to counter this restriction; however, SIVsm/SIVmac and HIV-2 express
Vpx to circumvent this restriction. Vpx targets SAMHD1 using the host cell E3 ubiquitin ligase complex, in which Vpx interacts with the DCAF1
subunit of the CUL4A/DDB1 ubiquitin ligase to degrade SAMHD1 via the proteasome. This allows HIV-1 reverse transcription to occur and viral
replication to complete.
St Gelais and Wu Retrovirology 2011, 8:55
http://www.retrovirology.com/content/8/1/55
Page 2 of 4

Vpx to overcome SAMHD1-mediated restriction, which
likely induces the innate antiviral immunity to confine
viral infection in natural hosts. Thus, the interactions
between SAMHD1 and Vpx may contribute to different
consequences of HIV-1 and HIV-2 infection in humans
(Table 1). Similarly, Manel and colleagues have sug-
gested that HIV-1 restriction in DCs allows HIV-1 to
avoid the antiviral immune responses derived from DCs,
which are critical antigen presenting cells bridging the
innate and adaptive immunity [12].
The biological function of SAMHD1 is largely
unknown. SAMHD1 mutations are involved in Aicardi-
Goutières syndrome (AGS), a genetic encephalopathy
mimicking congenital viral infection [13]. SAMHD1 was
initially cloned from human DCs as an interferon (IFN)-
g-inducible gene [14] and has been proposed to act as a
negative regulator of the IFN response [13]. The cellular
exonuclease TREX1 was recently shown to bind and
digest excess cytosolic HIV-1 DNA that would otherwise
activate type I IFN expression and trigger an innate
immune response [15]. Interestingly, similar to
SAMHD1,TREX1 mutations in humans are associated
with autoimmune and inflammatory diseases, including
AGS [15]. It is currently unknown whether polymorph-
isms of SAMHD1 and TREX1 are linked to AIDS pro-
gression or whether AGS patients are more susceptible
to HIV-1 infection.
Three major retrovirus restriction factors have been
identified: APOBEC3G, TRIM5a, and tetherin [6]. HIV-1
has developed mechanisms of evading these restriction
factors mainly through its accessory proteins, such as Vif
for APOBEC3G and Vpu for tetherin. These restriction
factors function across many different cell types, whereas
SAMHD1 appears to be specific to the myeloid-lineage
cells. It might be possible that SAMHD1 acts in concert
with another myeloid-specific co-factor [10]. It appears
that only Vpx from the SIVsm/HIV-2 lineage counteracts
SAMHD1-mediated HIV-1 restriction in myeloid cells
[9,10], while HIV-1 Vpr does not interact or degrade
SAMHD1 [10]. SAM domains are putative protein inter-
action modules that are capable of self-association and
binding to RNA and non-SAM domain containing pro-
teins. Given that SAMHD1 interferes with the accumula-
tion of HIV-1 reverse transcripts, one can speculate that
the SAM domain of SAMHD1 may bind HIV-1 RNA or
proteins and mediate their degradation through the HD
domain and the recruitment of the E3-ligase complex.
Further delineation of the mechanisms of SAMHD1
restriction is required to fully understand the HIV-1
restriction in myeloid-lineage cells and why HIV-1 has
not evolved a viral antagonist to counteract SAMHD1.
SAMHD1-mediated HIV-1 restriction has so far been
analyzed only in monocyte-derived DCs [9] and macro-
phages [10], and it should be investigated in primary
monocytes, myeloid DCs, as well as plasmacytoid DCs
that can produce high levels of type I IFN upon HIV-1
stimulation [1,2]. It is unclear whether SAMHD1 is also
type I IFN inducible, similar to other HIV-1 restriction
factors. It would be interesting to know whether
SAMHD1 can restrict other retroviruses, endogenous
retroviruses, or other non-retroviruses and whether
viruses use their own viral components to counteract
SAMHD1.
Conclusions
The discovery of SAMHD1 as a myeloid-cell-specific
HIV-1 restriction factor opens many intriguing ques-
tions in understanding intrinsic immunity against HIV-
1. When considering future therapeutic opportunities,
enhancement of SAMHD1 function may help hosts
develop potent innate and adaptive immune responses
to HIV-1. Further investigation of the mechanisms
underlying SAMHD1-mediated HIV-1 restriction will
shed light on the innate immune response against retro-
viruses and aid in the future development of more effec-
tive anti-HIV-1 interventions.
List of abbreviations
HIV-1: human immunodeficiency virus type 1; HIV-2: human
immunodeficiency virus type 2; SIV: simian immunodeficiency virus;
SAMHD1: sterile alpha motif domain- and HD domain-containing protein 1;
DDB1: damage-specific DNA binding protein 1; CUL4A: Cullin-4A; DCAF1:
DDB1- and CUL4A-associated factor-1; APOBEC3G: apolipoprotein B mRNA-
editing, enzyme-catalytic, polypeptide-like 3G; TRIM5α: tripartite motif-
containing protein 5α.
Acknowledgements
We thank members of the Wu laboratory for helpful discussions and critical
reading of the manuscript. The research in the Wu laboratory is supported
by grants (AI068493 and AI078762) to Li Wu from the NIH and by the
program of Public Health Preparedness for Infectious Diseases (PHPID) of
The Ohio State University. The authors apologize to all colleagues whose
work has not been cited as a result of space limitations.
Authors’contributions
Both authors contributed to the writing and editing of the manuscript and
approved the final manuscript.
Table 1 Comparison of HIV-1 and HIV-2 regarding
SAMHD1 degradation and potential disease
consequences.
Lentiviruses HIV-1 SIVsm/HIV-2
#
Vpx protein expression No Yes
Human SAMHD1 degradation No Yes
Efficient infection of myeloid cells No Yes
Triggering myeloid-cell-mediated
innate anti-viral immunity
through type I interferon
No Yes
Potential disease outcome Spread of
infection and
AIDS
Confined infection,
no AIDS in natural
hosts
#
It is known that cross-species transmission of sooty mangabey SIV (SIVsm) to
humans has given rise to HIV-2 [6].
St Gelais and Wu Retrovirology 2011, 8:55
http://www.retrovirology.com/content/8/1/55
Page 3 of 4

Competing interests
The authors declare that they have no competing interests.
Received: 21 June 2011 Accepted: 8 July 2011 Published: 8 July 2011
References
1. Wu L, KewalRamani VN: Dendritic-cell interactions with HIV: infection and
viral dissemination. Nat Rev Immunol 2006, 6:859-868.
2. Coleman CM, Wu L: HIV interactions with monocytes and dendritic cells:
viral latency and reservoirs. Retrovirology 2009, 6:51.
3. Goujon C, Jarrosson-Wuilleme L, Bernaud J, Rigal D, Darlix JL, Cimarelli A:
With a little help from a friend: increasing HIV transduction of
monocyte-derived dendritic cells with virion-like particles of SIV(MAC).
Gene Therapy 2006, 13:991-994.
4. Goujon C, Riviere L, Jarrosson-Wuilleme L, Bernaud J, Rigal D, Darlix JL,
Cimarelli A: SIVSM/HIV-2 Vpx proteins promote retroviral escape from a
proteasome-dependent restriction pathway present in human dendritic
cells. Retrovirology 2007, 4:2.
5. Ayinde D, Maudet C, Transy C, Margottin-Goguet F: Limelight on two HIV/
SIV accessory proteins in macrophage infection: is Vpx overshadowing
Vpr? Retrovirology 2010, 7:35.
6. Malim MH, Emerman M: HIV-1 accessory proteins–ensuring viral survival
in a hostile environment. Cell Host Microbe 2008, 3:388-398.
7. Gibbs JS, Lackner AA, Lang SM, Simon MA, Sehgal PK, Daniel MD,
Desrosiers RC: Progression to AIDS in the absence of a gene for vpr or
vpx. J Virol 1995, 69:2378-2383.
8. Hirsch VM, Sharkey ME, Brown CR, Brichacek B, Goldstein S, Wakefield J,
Byrum R, Elkins WR, Hahn BH, Lifson JD, Stevenson M: Vpx is required for
dissemination and pathogenesis of SIV(SM) PBj: evidence of
macrophage-dependent viral amplification. Nat Med 1998, 4:1401-1408.
9. Laguette N, Sobhian B, Casartelli N, Ringeard M, Chable-Bessia C, Segeral E,
Yatim A, Emiliani S, Schwartz O, Benkirane M: SAMHD1 is the dendritic-
and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx.
Nature 2011, 474:654-657.
10. Hrecka K, Hao C, Gierszewska M, Swanson SK, Kesik-Brodacka M,
Srivastava S, Florens L, Washburn MP, Skowronski J: Vpx relieves inhibition
of HIV-1 infection of macrophages mediated by the SAMHD1 protein.
Nature 2011, 474:658-661.
11. Zimmerman MD, Proudfoot M, Yakunin A, Minor W: Structural insight into
the mechanism of substrate specificity and catalytic activity of an
HD-domain phosphohydrolase: the 5’-deoxyribonucleotidase YfbR from
Escherichia coli. J Mol Biol 2008, 378:215-226.
12. Manel N, Hogstad B, Wang Y, Levy DE, Unutmaz D, Littman DR: A cryptic
sensor for HIV-1 activates antiviral innate immunity in dendritic cells.
Nature 2010, 467:214-217.
13. Rice GI, Bond J, Asipu A, Brunette RL, Manfield IW, Carr IM, Fuller JC,
Jackson RM, Lamb T, Briggs TA, et al:Mutations involved in Aicardi-
Goutieres syndrome implicate SAMHD1 as regulator of the innate
immune response. Nat Genet 2009, 41:829-832.
14. Li N, Zhang W, Cao X: Identification of human homologue of mouse
IFN-gamma induced protein from human dendritic cells. Immunol Lett
2000, 74:221-224.
15. Yan N, Regalado-Magdos AD, Stiggelbout B, Lee-Kirsch MA, Lieberman J:
The cytosolic exonuclease TREX1 inhibits the innate immune response to
human immunodeficiency virus type 1. Nat Immunol 2010, 11:1005-1013.
doi:10.1186/1742-4690-8-55
Cite this article as: St Gelais and Wu: SAMHD1: a new insight into HIV-1
restriction in myeloid cells. Retrovirology 2011 8:55. Submit your next manuscript to BioMed Central
and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at
www.biomedcentral.com/submit
St Gelais and Wu Retrovirology 2011, 8:55
http://www.retrovirology.com/content/8/1/55
Page 4 of 4



![PET/CT trong ung thư phổi: Báo cáo [Năm]](https://cdn.tailieu.vn/images/document/thumbnail/2024/20240705/sanhobien01/135x160/8121720150427.jpg)






















%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)