
THE EMBO LECTURE
Diversity of human U2AF splicing factors
Based on the EMBO Lecture delivered on 7 July 2005 at the
30th FEBS Congress in Budapest
Ine
ˆs Mollet, Nuno L. Barbosa-Morais, Jorge Andrade and Maria Carmo-Fonseca
Instituto de Medicina Molecular, Faculdade de Medicina, Universidade de Lisboa, Portugal
Introduction
In eukaryotes, protein-coding regions (exons) within
precursor mRNAs (pre-mRNAs) are separated by
intervening sequences (introns) that must be removed
to produce a functional mRNA. Pre-mRNA splicing is
an essential step for gene expression, and the vast
majority of human genes comprise multiple exons that
are alternatively spliced [1]. Alternative splicing is used
to generate multiple proteins from a single gene, thus
contributing to increase proteome diversity. Alternative
splicing can also regulate gene expression by generating
mRNAs targeted for degradation [2]. Proteins
produced by alternative splicing control many physio-
logical processes and defects in splicing have been
linked to an increasing number of human diseases [1,3].
Pre-mRNA splicing occurs in a large, dynamic com-
plex called the spliceosome. The spliceosome is com-
posed of small nuclear ribonucleoprotein particles (the
U1, U2, U4 ⁄U5 ⁄U6 snRNPs forming the major
spliceosome and the U11, U12, U4atac ⁄U6atac.U5
snRNPs forming the less abundant minor spliceosome)
and more than 100 non-snRNP proteins [4]. Spliceo-
some assembly follows an ordered sequence of events
that begins with recognition of the 5¢splice site by
U1snRNP and binding of U2AF (U2 small nuclear
ribonucleoprotein auxiliary factor) to the polypyrimi-
dine (Py)-tract and 3¢splice site [5]. Human U2AF is a
heterodimer composed of a 65-kDa subunit (U2AF
65
),
which contacts the Py-tract [6–8], and a 35-kDa sub-
unit (U2AF
35
), which interacts with the AG dinucleo-
tide at the 3¢splice site [9–11]. Assembly of U2AF
with the pre-mRNA, which in yeast and mammals
requires an interaction with the U1 snRNP [12–17], is
important for subsequent recruitment of U2snRNP to
the spliceosome.
U2AF has been highly conserved during evolution.
In addition, a number of U2AF-related genes are
Keywords
CAPER; PUF60; RNA splicing; U2AF
Correspondence
M. Carmo-Fonseca, Institute of Molecular
Medicine, Faculty of Medicine, Avenue Prof.
Egas Moniz, 1649–028 Lisbon, Portugal
Fax: +351 21 7999412
Tel: +351 21 7999411
E-mail: carmo.fonseca@fm.ul.pt
(Received 13 July 2006, revised 12 Septem-
ber 2006, accepted 14 September 2006)
doi:10.1111/j.1742-4658.2006.05502.x
U2 snRNP auxiliary factor (U2AF) is an essential heterodimeric splicing
factor composed of two subunits, U2AF
65
and U2AF
35
. During the past
few years, a number of proteins related to both U2AF
65
and U2AF
35
have
been discovered. Here, we review the conserved structural features that
characterize the U2AF protein families and their evolutionary emergence.
We perform a comprehensive database search designed to identify U2AF
protein isoforms produced by alternative splicing, and we discuss the
potential implications of U2AF protein diversity for splicing regulation.
Abbreviations
EST, expressed sequence tag; FIR, FUSE-binding protein-interacting repressor; PUF60, poly(U)-binding factor-60 kDa; RRM, RNA-recognition
motif; SF1, splicing factor 1; U2AF, U2 small nuclear ribonucleoprotein auxiliary factor; UHM, U2AF homology motif.
FEBS Journal 273 (2006) 4807–4816 ª2006 The Authors Journal compilation ª2006 FEBS 4807

present in the human genome, and some are known to
be alternatively spliced. Here, we review currently
available information on the diversity of U2AF pro-
teins and we discuss the resulting implications for
splicing regulation.
Structural features of U2AF and
U2AF-related proteins
The U2AF
65
protein contains three RNA-recognition
motifs or RRMs (Table 1). The two central motifs
(RRM1 and RRM2) are canonical RRM domains
responsible for recognition of the Py-tract in the pre-
mRNA, whereas the third RRM has unusual features
and is specialized in protein–protein interaction. This
unusual RRM-like domain, called UHM for U2AF
homology motif, is present in many other splicing pro-
teins [18]. The UHM in U2AF
65
recognizes splicing
factor 1 (SF1), and this cooperative protein–protein
interaction strengthens the binding to the Py-tract
(Fig. 1). The UHM motif was highly conserved from
yeast to mammals, but, paradoxically, appears dispen-
sable for splicing of at least certain pre-mRNAs
in vitro [19]. The N-terminal amino acids 85–112 of
U2AF
65
interact with U2AF
35
, and this association
further strengthens the binding to the Py-tract [18].
Although it is not a member of the serine-arginine
(SR) family of splicing factors, the U2AF
65
protein
further contains an arginine and serine rich (RS)
domain that is required for spliceosome assembly
in vitro [20,21]. Importantly, binding of U2AF
65
alone
is sufficient to bend the Py-tract, juxtaposing the
branch region and 3¢splice site [22]. Current models
therefore propose an arrangement in which the
C-terminus of U2AF
65
is positioned proximal to the
branch point, and the N-terminus is situated in
the vicinity of the 3¢splice site (Fig. 1).
PUF60 [poly(U)-binding factor-60 kDa] was first
isolated as a protein closely related to U2AF
65
that
was required for efficient reconstitution of RNA spli-
cing in vitro [23]. The homology between PUF60 and
U2AF
65
extends across their entire length, except for
the N-terminus where PUF60 lacks a recognizable
RS domain (Table 1 and Fig. 2A). CAPERaand
CAPERbare the most recently characterized proteins
related to U2AF
65
[24]. Both have a domain organiza-
tion similar to U2AF
65
, except for the C-terminus of
CAPERbwhich lacks the UHM domain (Table 1 and
Fig. 2A).
The U2AF
35
protein contains a central UHM
domain (previously called Y-RRM) involved in the
interaction with U2AF
65
, flanked by two Zn
2+
-binding
motifs and a C-terminal RS domain (Table 2 and
Fig. 1). Three-dimensional structural information
revealed that, despite low primary sequence identity
(23%), recognition of the respective ligands by the
U2AF
65
-UHM and U2AF
35
-UHM domains is very
similar [18]. Both the U2AF
35
–U2AF
65
and U2AF
65
–
SF1 interactions involve a critical Trp residue in the
ligand sequence which inserts into a tight hydrophobic
pocket created by the UHM (Fig. 3).
In the human genome there are at least three genes
that encode proteins with a high degree of homology
to U2AF
35
(Table 2 and Fig. 2B). U2AF
26
(encoded
by the U2AF1L4 gene) is a 26-kDa protein bearing
strong sequence similarity to U2AF
35
; the N-terminal
187 amino acids are 89% identical, but the C-terminus
of U2AF
26
lacks the RS domain present in U2AF
35
[25]. U2AF
35
R1 (encoded by the U2AF1L1 gene) and
Table 1. Domain organization of U2AF
65
and U2AF
65
-related pro-
teins. Domains are annotated as described in [18]. RS, Arg-Ser rich.
The gene names approved by the HUGO Gene Nomenclature Com-
mittee (http://www.gene.ucl.ac.uk/nomenclature/) have been inclu-
ded.
Gene Protein Domain organization
U2AF2 U2AF
65
475aa
SIAHBP1 PUF60 559aa
RNPC2 CAPERa530aa
RBM23 CAPERb424aa
SF1
U2AF65
U2AF35
5’
Fig. 1. Schematic representation of protein–protein and protein–RNA
interactions mediated by the U2AF heterodimer during the early
steps of spliceosome assembly. Binding of the U2AF heterodimer to
the Py-tract and 3¢-splice site AG is strengthened by the co-operative
interaction between U2AF
65
and SF1 at the branchpoint (encircled A)
sequence (BPS). Binding of U2AF
65
bends the Py-tract (solid line) to
bring the 3¢splice site and BPS region close together. The ligand Trp
residues (W) in SF1 and U2AF
65
insert into the UHM pockets in
U2AF
65
and U2AF
35
, respectively. An additionally exposed Trp resi-
due on the U2AF
35
UHM domain inserts between a series of unique
Pro residues at the N-terminus of U2AF
65
(P).
U2AF diversity I. Mollet et al.
4808 FEBS Journal 273 (2006) 4807–4816 ª2006 The Authors Journal compilation ª2006 FEBS

U2AF
35
R2 ⁄Urp (encoded by the U2AF1L2 gene) are
94% identical with one another and contain stretches
that are 50% identical to corresponding regions of
U2AF
35
[26]. Additional sequences encoding putative
new proteins related to U2AF
35
have been identified in
the human genome [27,28], but these have not yet been
characterized experimentally.
Evolution of U2AF genes
Phylogenetic analysis indicates that the origin of
U2AF gene families dates back to the divergence of
the eukaryotes, more than 1500 million years ago [28].
Orthologs of both U2AF
65
and U2AF
35
are found in
Drosophila melanogaster [29,30], Caenorhabditis elegans
[10,31], Schizosaccharomyces pombe [32,33], Arabidop-
sis thaliana [34], and Plasmodium falciparum [28]. In
contrast, the genome of Saccharomyces cerevisiae con-
tains a poorly conserved ortholog of the U2AF large
subunit, Mud2p, and no open reading frame that
resembles the small subunit [35]. Orthologs of human
PUF60 are present across metazoans, while CAPER
proteins are found all across the eukaryotic lineage.
Orthologs of U2AF
35
R2 ⁄Urp exist in insects, chor-
dates and vertebrates (Fig. 4).
Phylogenetic studies show that both the U2AF
35
and CAPER genes were most likely duplicated during
the wave of whole-genome duplications that occurred
at the early emergence of vertebrates 650–450 million
years ago, giving rise to U2AF
26
and CAPERb,
respectively. Orthologs of either U2AF
26
or CAPERb
are not detected in lower eukaryotes such as Dro-
sophila,C. elegans or plants. Intriguingly, these two
genes were apparently lost in some vertebrate lineages
and remained in others (Fig. 4). Orthologs of U2AF
26
are present in the human and mouse genomes, and
expressed sequence tags (ESTs) more similar to
U2AF
26
than U2AF
35
are found in rat, pig, and cow.
However, there is no evidence for the existence of the
gene encoding U2AF
26
in the genomes of birds,
amphibians or fish. A comparison of the mouse and
human U2AF1L4 gene revealed that the exon ⁄intron
boundaries are located in the same positions as in the
human U2AF1 gene, although the introns are much
U2AF65
U2AF35
U2AF26
U2AF35R1
U2AF35R2
PUF60
CAPERα
CAPERβ
Fig. 2. A schematic alignment of human
proteins related to U2AF
65
(A) and U2AF
35
(B). (A) The putative functional domains in
each protein are aligned with U2AF
65
, and
the similarity (% identity) of these domains
in relation to U2AF
65
is indicated. (B) The
putative functional domains in each protein
are aligned with U2AF
35
, and the similarity
(% identity) of these domains in relation to
U2AF
35
is indicated.
Table 2. Domain organization of U2AF
35
and U2AF
35
-related
proteins. Domains are annotated as described in [18]. Zn, zinc
binding; RS, Arg-Ser rich. The gene names approved by the HUGO
Gene Nomenclature Committee (http://www.gene.ucl.ac.uk/
nomenclature/) have been included.
Gene Protein Domain organization
U2AF1 U2AF
35
240aa
U2AF1L4 U2AF
26
202aa
U2AF1L1 U2AF
35
R1 479aa
U2AF1L2 U2AF
35
R2 482aa
I. Mollet et al.U2AF diversity
FEBS Journal 273 (2006) 4807–4816 ª2006 The Authors Journal compilation ª2006 FEBS 4809
smaller in the U2AF1L4 gene. In addition, the exon
sequences of the human and mouse U2AF1L4 genes
are 90% identical at the nucleotide level, and the
majority of the differences are neutral, third-position
changes [25]. The evolutionary pattern for CAPERbis
more unusual. Among mammals, orthologs can be
found for primates (chimp and rhesus) and domestic
animals (dog and cow) but not for rodents. CAPERb
can also be found in Xenopus tropicalis, but there is no
evidence for its existence in chicken or fish. A compar-
ison of CAPERbgenes from different mammals
revealed that most of the exon ⁄intron boundaries are
located in the same positions as in the human
CAPERagene and the introns are found to be smaller
in the CAPERbgene. Given the similarities between
the evolutionary histories of the U2AF
26
and CAPERb
genes, it is likely that these new splicing proteins per-
form unique and lineage-specific functions.
Retrotransposition rather than gene duplication
appears to have created the U2AF1L1 gene less than
100 million years ago. The mouse U2AF1L1 gene,
which is located on chromosome 11, was formed by
retrotransposition of U2AF1L2, which is located on
the X chromosome [36]. U2AF1L1 is regulated by
genomic imprinting [37], and the whole gene is located
in an intron of another gene, Murr1, that is not
imprinted [36]. The retrotransposition that originated
the mouse U2AF1L1 gene must have occurred after
mice and humans diverged, because the human ortho-
log of Murr1 is located on chromosome 2 and there
are no U2AF1-related genes on human chromosome 2.
Indeed, the phylogenetic analysis of this family of
genes indicates independent events of retrotrans-
position in rodents (mouse and rat) and primates
(human and chimp). Similarly to the mouse gene, the
human U2AF1L1 gene located on chromosome 5 is
intronless whereas human U2AF1L2 is multiexonic,
suggesting that it also originated by retrotransposition
[28]. However, in contrast with the mouse gene, human
U2AF1L1 is not imprinted [38].
Alternative splicing and diversity of
human U2AF proteins
Our laboratory has recently reported that human tran-
scripts encoding U2AF
35
can be alternatively spliced
giving rise to three different mRNA isoforms called
U2AF
35
a, U2AF
35
b, and U2AF
35
c [39]. This discovery
raised the question of whether additional U2AF genes
produce alternatively spliced mRNAs. Very few
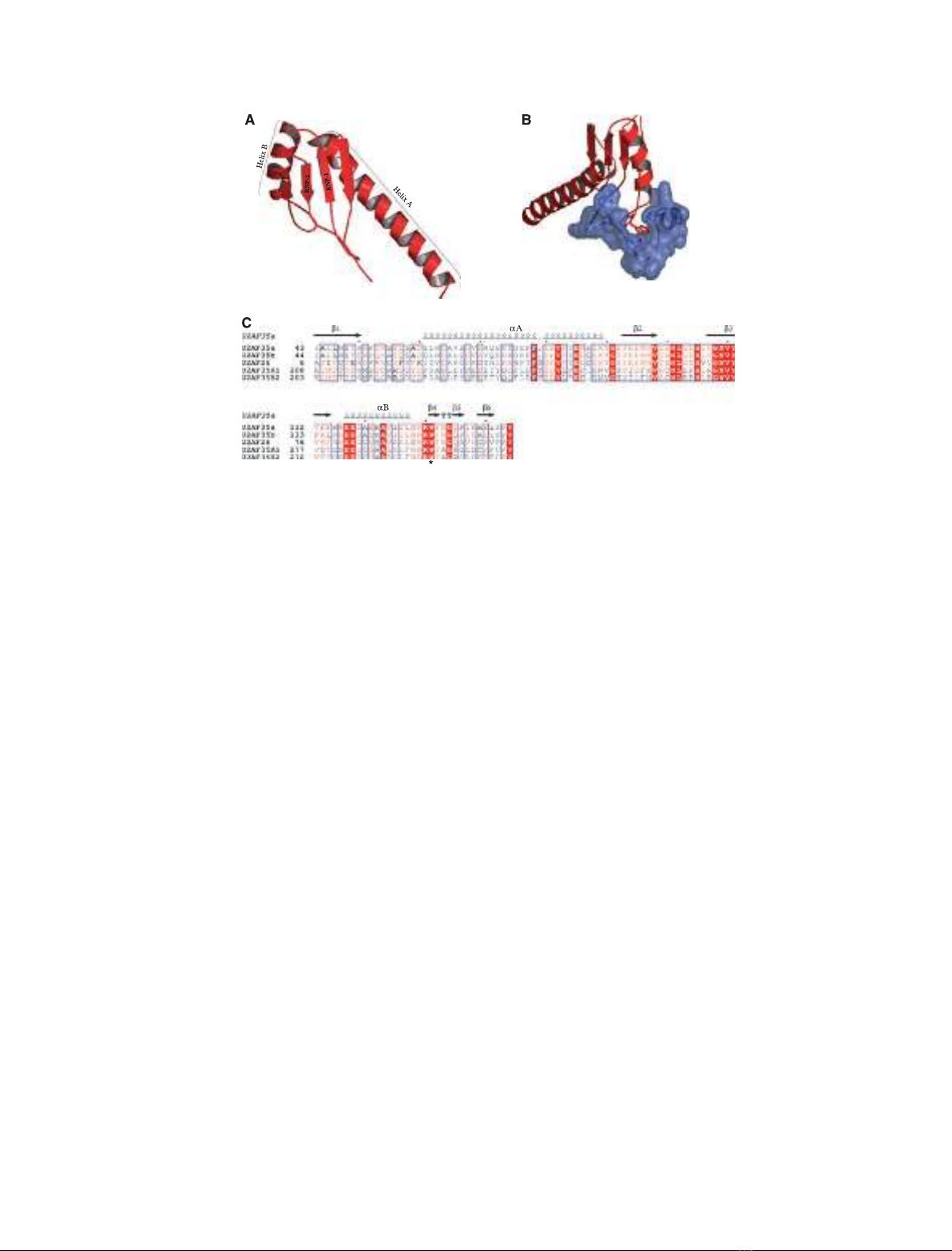
Fig. 3. (A) Ribbon representation of the U2AF
35
UHM. Residues 43–146; pdb code: 1jmt. (B) Structure of the U2AF
35
UHM (red)–U2AF
65
lig-
and (blue) complex [64]. A critical W residue (Trp92 in U2AF
65
) inserts into a tight hydrophobic pocket between the a-helices and the RNP1-
and RNP2-like motifs in U2AF
35
[64]. An Arg residue (Arg133 in U2AF
35
) on the loop connecting the last a-helix and b-strand of the UHM
contributes to the Trp-binding pocket. A neighboring W residue (Trp134 in U2AF
35
) inserts between a series of unique Pro residues at the
N-teminus of U2AF
65
(residues 85–112). In addition, a series of acidic residues in helix A of the UHM interacts with basic residues at
the N-terminus of U2AF
65
. The molecular representations were generated using PYMOL [65]. (C) Sequence alignment of the UHM region in
the alternatively spliced U2AF
35
isoforms (U2AF
35
a and U2AF
35
b) and in the genes that encode U2AF
35
-related proteins. The conserved Trp
residues are identified by an asterisk. The alignment was generated by the program MULTALIN [66], and the figure was prepared using ESPRIPT
[67]. The secondary structure of U2AF
35
, derived from 3D data [64], is represented in the upper part of the alignment.
U2AF diversity I. Mollet et al.
4810 FEBS Journal 273 (2006) 4807–4816 ª2006 The Authors Journal compilation ª2006 FEBS
examples of U2AF mRNA isoforms have been des-
cribed in the literature. Namely, two CAPERb
mRNAs and four CAPERamRNAs were detected in
several human tissues by northern blotting [24], and a
splicing variant of PUF60 ⁄FIR was identified in colo-
rectal cancers [40]. This scarcity of data prompted us
to use bioinformatic search strategies to investigate
alternative splicing of U2AF and U2AF-related genes.
This analysis was carried out with the aid of the
UCSC Genome Browser (http://genome.ucsc.edu/) [41]
for the human genome assembly hg17, May2004,
NCBI Build 35. Gene regions of interest were defined
by the BLAT mapping [41] of the available RefSeq
transcript (RNA) sequences [42] (http://www.ncbi.nlm.
nih.gov/projects/RefSeq/) for a particular gene. Using
the UCSC Table Browser [43], we obtained the tables
for the BLAT mappings of mRNAs and ESTs for this
gene region. Making allowance only for GT_AG,
GC_AG or AT_AC splice site consensus and excluding
isoforms with extensive intron retentions, the non-
redundant set of longest isoforms and corresponding
accessions was determined. The splicing patterns
obtained were cross-checked with two alternative spli-
cing databases: the ASAP (http://bioinfo.mbi.ucla.edu/
ASAP/); and the Hollywood RNA Alternative Splicing
Database (http://hollywood.mit.edu).
Our analysis revealed that, with the single exception
of the U2AF1L1 gene, which is devoid of introns, all
genes coding for U2AF and U2AF-related proteins
can be alternatively spliced (Table 3). Many alternat-
ively spliced mRNA isoforms are predicted to contain
premature stop codons and are therefore expected to
be targeted for degradation by nonsense-mediated
decay, as already demonstrated for U2AF
35
c (corres-
ponding to RefSeq mRNA NM_001025204 in
Table 3). In addition, we found evidence for several
transcripts that could generate functional protein iso-
forms containing the conserved RRM motifs charac-
teristic of each protein family (Table 3). Variations in
activity are expected from changes in domain structure
predicted for some of these isoforms, but further
experimental studies are needed to address this view.
Perspectives: evolution of U2AF
functions
After the discovery that U2AF
65
is required to recons-
titute mammalian splicing in vitro [6–8], the protein
FA2U
53
FA2U
62
F
A2U
53
1
R
FA2U
53
2R
FA2U
5
6
06FUP
REPAC α
R
E
PA
Cβ
0
0
0
5
00
010
0
51
ayM
rtort
ern
oitis
o
p
s
na s ni oe
m
ma
mmila al
ninese
g
a
noit
aci
lpu
d
em
o
neg
elohw
inr
ay-finh
s
i
f
d
e
n
1-one
g elohw 2 med,
snoit
acilpu
rev tecnegrevid
e
t
a
rbe
stso
e
let ni detacilpud
prto z
oao
ey ts
as
r
o
ws
m
st
c
esni
ca
noi
h
si
f
stn
e
dor
ecnegrevid
n
am
u
h m
or
f
p
m
an
aib
ih s
s
dri
b
de
m
ot
sci n
a .
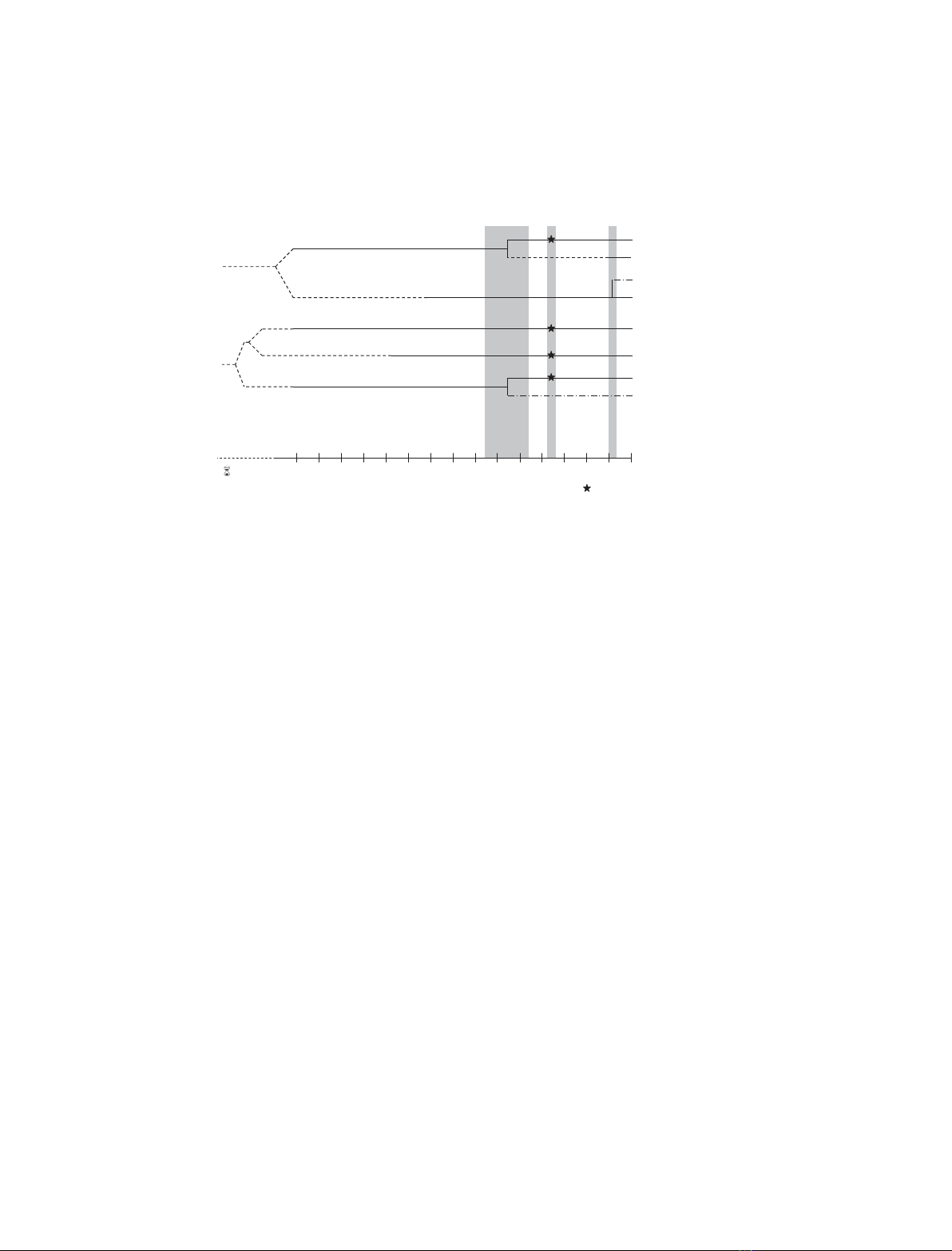
Fig. 4. Evolution of U2AF-related proteins. The possible origins of U2AF proteins are shown in relation to key metazoan evolutionary events.
Solid lines represent presence of the indicated protein in all species that diverged from humans within the corresponding period of time.
Dashed lines represent loss of the indicated proteins in all extant species that diverged from humans within the corresponding period of
time. Dashed-dotted lines represent lineage-specific loss ⁄preservation or appearance ⁄absence of the indicated protein in species that
diverged from humans within the corresponding period of time (e.g. CAPERbapparently disappeared from fish, birds and rodents but
remained in Xenopus and some mammals; U2AF
35
R1 results from independent retrotransposition events affecting only primates and
rodents). A star indicates that U2AF
35
, U2AF
65
, PUF60 and CAPERagenes are duplicated in teleosts, most probably as a consequence of
the whole-genome duplication that occurred in ray-finned fish 350 million years ago (Mya).
I. Mollet et al.U2AF diversity
FEBS Journal 273 (2006) 4807–4816 ª2006 The Authors Journal compilation ª2006 FEBS 4811


























%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)