
Di truyền học quần thể ( phần 1 )
Trạng thái cân bằng của quần thể giao phối
1. Định luậtHardy-weinberg
Năm 1908, nhà toán học người Anh G.N. Hardy và một bác sĩ người Đức
là W. Weinberg đã độc lập nghiên cứu, đồng thời phát hiện ra quy luật
phân bố các kiểu gen và kiểu hình trong quầnthể giao phối.
Nội dung định luật chỉ ra rằng, trongnhững điều kiện nhất định, thì trong
lòng một quần thể giao phối tần số tương đối của các alen ở mỗi gen có
khuynh hướng duy trì không đổi từ thế hệ nay sang thế hệ khác.
Ví dụ trong trường hợp đơn giản, gen A có 2 alen là A và a, thì trong
quần thể có 3 kiểu trên AA, Aa, aa. Giả sử các kiểu gen trong quần thể ở
thế hệ xuất phát là:
0,64AA + 0,32Aa + 0,04aa = 1 thì tần số tương đối củaalen A là: và tần
số tươngđốicủaalen a là: 0,04 + 0,32/2 = 0,2
Tần số của A so với a ở thế hệ xuất phát là: 0,64 + 0,32/2 = 0,8
A/a = 0,8/0,2. Tỷ lệ này có nghĩa là trong số các giao tử đực cũng như
giao tử cái, số giao tử mang alen A chiếm 80%, còn sốgiao tử mang alen
a chiếm 20%. Khi tổ hợp tử do các loại giao tử này tạo ra thế hệ tiếptheo
như sau:
♂
♀
0,8A 0,2a
0,8A 0,64AA 0,16Aa
0,2a 0,16Aa 0,04aa
Tần số tương đối các kiểu gen ở thế hệ này là: 0,64AA + 0,32Aa + 0,04aa
= 1 và tần sốtương đối các alen: A/a = 0,8/0,2
Trong các thế hệ tiếp theo tần số đó vẫn không thay đổi.
Simpo PDF Merge and Split Unregistered Version - http://www.simpopdf.com

Tổng quát: Nếu một gen A có 2 alen: A với tần số p; a với tần số q thì ta
có công thức: P2AA + 2pqAa + q2 aa = 1 .
Lưuý rằng: p2 + 2pq + q2 = (p + q)2 = 1 => p + q = 1 => q = (1 - p)
Do đó sự phân bố các kiểu trên còn có thể diễnđạt bằng: [pA + (1 - p) a]2
= 1
Nếu gen A có số alen nhiều hơn 2, ví dụ: a1, a2, a3... với các tần số tương
ứng p,
q, r,... thì sự phân bố các kiểu trên trong quần thể sẽ tương ứng với sự
triển khai biểu thức(p + q + r +...)2 =1.
Định luật Hardy-weinberg chỉ áp dụng cho quần thể giao phối và chỉ
nghiệm đúng trongnhững điều kiện sau:
- Có sự giao phối tự do, nghĩa là cá cá thể có kiểu gen và kiểu hình khác
nhau trong quần thể đều giao phối được với nhau, với xác suất ngang
nhau. Đây là điều kiện cơ bản nhất.
- Quần thể phảicó số lượng cá thể đủ lớn.
- Các loại giao tử mang alen trội, lặn được hình thành qua giảm phân với
tỷ lệ ngang nhau, có sức sống nhưnhau, tham gia vào thụ tinh vớixác suất
ngang nhau.
- Các cơ thể đồng hợp và dị hợp có sức sống ngang nhau, được truyền
gen cho các thếhệsau ngang nhau.
- Không có áp lựccủaquá trình đột biến và quá trình chọn lọctự nhiên
hoặc áp lực đó là không đángkể.
- Quần thể đượccách ly với các quần thể khác,không có sự trao đổi gen.
2. Ý nghĩa địnhluậtHardy-weinberg
Về thực tiễn, dựa vào công thức Hardy-weinberg có thể từ tỷ lệ kiểu hình
suy ra tỷ lệ kiểu gen và tần số tương đối các alen, ngược lại, từ tần số
tương đối của alen đãbiết có thể dự tính tỷ lệ các kiểugen và kiểu hình.
Nắm được kiểu gen của một số quần thể có thể dự đoán tác hại của các
đột biến gây chết, đột biến có hại, hoặc khả năng gặp những đồnghợptử
mang đột biếncó lợi.
Simpo PDF Merge and Split Unregistered Version - http://www.simpopdf.com
Về lý luận, định luật Hardy-weinberg giải thích vì sao trong tự nhiên có
những quần thể đứng vững trạng thái ổn định trong thời gian dài. Trong
tiến hoá, sự duy trì,kiên định những đặc điểm đạt được có ý nghĩa quan
trọng chứ không phải chỉ có sự phát sinh các đặc điểm mới mới có ý
nghĩa.
Cấu trúc di truyền của quần thể
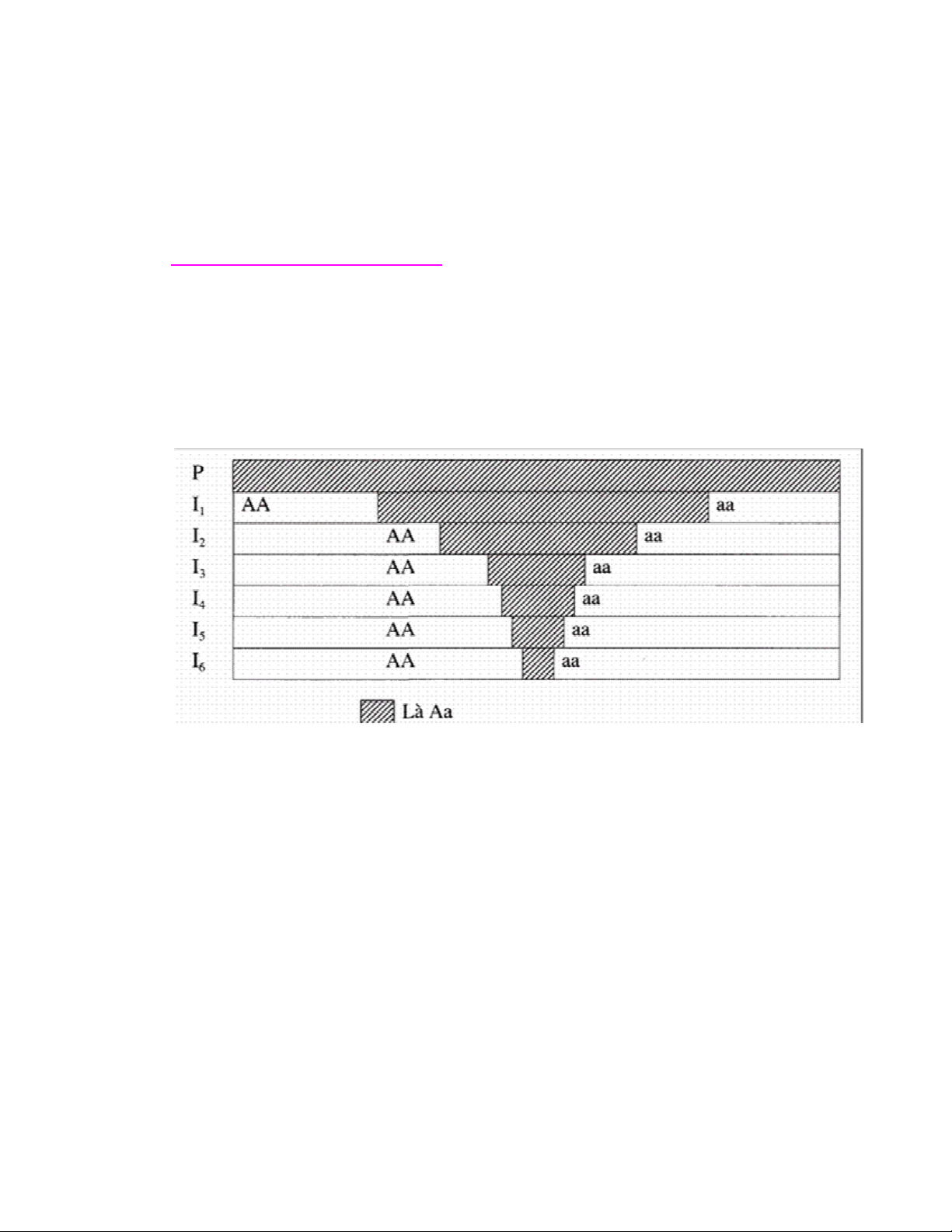
1. Quần thể tự phối
Tự phối hay giao phối gần (gọi chung là nội phối) làm cho quần thể dần
dần bị phân thành những dòng thuần có kiểu gen khác nhau. Trải qua
nhiều thế hệ nội phối, các gen ở trạng thái dị hợp chuyển sang trạng thái
đồng hợp. Số thể dị hợp giảm dần, số đồng hợp tăng dần.
Sơ đồ 1: Sự biến đổi cấu trúc di truyền của quần thể tự phối qua các thế
hệ
2. Quần thể giao phối
a/ Tính đa hình của quần thể giao phối
Quá trình giao phối là nguyên nhân làm cho quần thể đa hình về kiểu
trên, sự đa hình về kiểu trên tạo nên sự đa hình về kiểu hình.
Chẳng hạn, một gen A có a alen a1 và a2 qua giao phối tự do ra 3 tổ hợp
a1a1, a2a2. Nếu gen A có 3 alen a1, a2, a3 sẽ tạo ra 6 tổ hợp a1a1, a1a2, a1a3,
a2a2, a2a3, a3a3. Tổng quát, nếu gen A có r alen thì qua giao phối tự do, số
tổ hợp về gen A sẽ là: GA = r(r -1)/2
Simpo PDF Merge and Split Unregistered Version - http://www.simpopdf.com

Nếu có 2 gen A và B nằm trên những nhiễm sắc thể khác nhau, thì số tổ
hợp các alen về cả 2 gen A và B cùng một lúc sẽ là: G = g A x g B
Ví dụ gen A có 3 alen, gen B có 4 alen, thì G = 6 x 10 = 60.
Tất cả các tổ hợp gen trong quần thể tạo nên vốn gen (gen pool) của quần
thể đó.
Sự đa hình về kiểu trên bao giờ cũng phong phú hơn sự đa hình về kiểu
hình, vì sự biểu hiện kiểu hình của một alen đòi hỏi những tổ hợp đen xác
định và điều kiện ngoại cảnh thuận lợi. Một quần thể được gọi là đa hình
khi trong quần thể tồn tại nhiều kiểu hình khác nhau ở trạng thái cân bằng
tương đối ổn định. Trong một quần thể có thể đa hình về tính trạng này
nhưng đơn hình về tính trạng khác.
b/ Tần số tương đối của các alen trong quần thể giao phối
Tỷ lệ phần trăm mỗi loại kiểu hình trong quần thể được gọi là tần số
tương đối các kiểu hình.
Tần số tương đối của một hiện được tính bằng tỷ lệ phần trăm số giao tử
mang alen đó. Từ tỷ lệ phân bố các kiểu hình có thể suy ra tỷ lệ phân bố
các kiểu trên và từ đó suy ra tần số tương đối của các diễn. Ví dụ: có thể
tính tần số tương đối của các diễn M và N (hệ nhóm máu M, N) trong hai
quần thể người như sau:
Bảng 1 : Tần số tương đối của các alen M và N
Kiểu
hình:
M
N
N
Kiểu
gen:
M
N
N
N
Tần số tương
đối
của alen
Quầ
n thể
Số
cá
th
ể
được
NC
Số
lượn
g
% Số
lượng
% Số
lượng
%
M
N
1.Da
trắn
g ch.
6.12
9
1.787
29,1
6
3.039 49,58 1.303 21,26 0,539 0,460
Simpo PDF Merge and Split Unregistered Version - http://www.simpopdf.com

2.Thổ
dân Úc
730 22 3,01
216 29,59 492 67,40 01178
0,822
Những người thuộc nhóm máu M có kiểu trên MM cho ra toàn giao tử
mang gen M, những người thuộc nhóm máu N, có kiểu gen NN, cho ra
toàn giao tử mang men N, những người thuộc nhóm máu MN, có kiểu
trên MN, cho một nửa giao tử mang alen M và một nửa số giao tử mang
alen N.
Tần số tương đối alen M ở quần thể 2 là: 3.01% + (29,59%)/2 + 17,79%
= 0,178%
Điều này có nghĩa, trong quần thể này cứ 1000 giao tử thì có 178 giao tử
mang alen M, còn 822 giao tử mang alen N. Tần số này thay đổi tuỳ quần
thể người.
Tóm lại, tần số tương đối của các alen về một bên nào đó là dấu hiệu đặc
trưng cho sự phân bố kiểu gen và kiểu hình trong quần thể đó.
Chọn lọc tự nhiên tác động lên thành phần di truyền của quần thể
Cả ba quá trình gây biến đổi tần số gene đã xét trên đây đều có một điểm
chung là không một quá trình nào định hướng đối với sự thích nghi
(adaptation). Theo nhận định của một số tác giả (Mayer 1974; Ayala và
Kiger 1980), các quá trình này là ngẫu nhiên đối với sự thích nghi, do đó
tự thân chúng sẽ phá hoại tổ chức và các đặc tính thích nghi của sinh vật.
Chỉ có chọn lọc tự nhiên (natural selection) mới là quá trình thúc đẩy sự
thích nghi và hạn chế các hiệu quả phá hoại tổ chức của các quá trình
khác. Trong ý nghĩa đó, chọn lọc tự nhiên là quá trình tiến hóa khốc liệt
nhất, bởi vì chỉ có nó mới có thể giải thích được bản chất thích nghi, tính
đa dạng (diversity) và có tổ chức cao của các sinh vật.
Ý tưởng về chọn lọc tự nhiên như là quá trình nền tảng, là động lực của
sự biến đổi tiến hóa do Charles Darwin và Alfred Russel Wallace độc lập
đưa ra năm 1858. Lý luận tiến hóa bằng chọn lọc tự nhiên đã được phát
triển đầy đủ, với chứng cứ ủng hộ xác đáng, trong cuốn Nguồn gôc các
loài (The Origin of Species) do Darwin xuất bản năm 1859.
Theo Hartl et al (1988, 1997), trên quan điểm thuyết tổng hợp hiện đại,
có thể hình dung chọn lọc tự nhiên xảy ra dựa trên ba điểm chính: (1) Ở
mọi sinh vật, đời con được sinh ra nhiều hơn số sống sót và sinh sản; (2)
Các cá thể khác nhau về khả năng sống sót và sinh sản và phần lớn những
Simpo PDF Merge and Split Unregistered Version - http://www.simpopdf.com

![Hướng dẫn tư vấn di truyền học sinh sản cho bệnh nhân [chuẩn nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2018/20180728/trieuhkt/135x160/8131532770629.jpg)



![Đột biến số lượng nhiễm sắc thể: Chuyên đề [Chuẩn SEO]](https://cdn.tailieu.vn/images/document/thumbnail/2016/20161024/uceacademy/135x160/5091477305546.jpg)




















%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)