
6. B gen ti thộ ể
6.1. B gen ti th các loài sinh v t (Tr nh Ng c Đc)ộ ể ở ậ ị ọ ứ
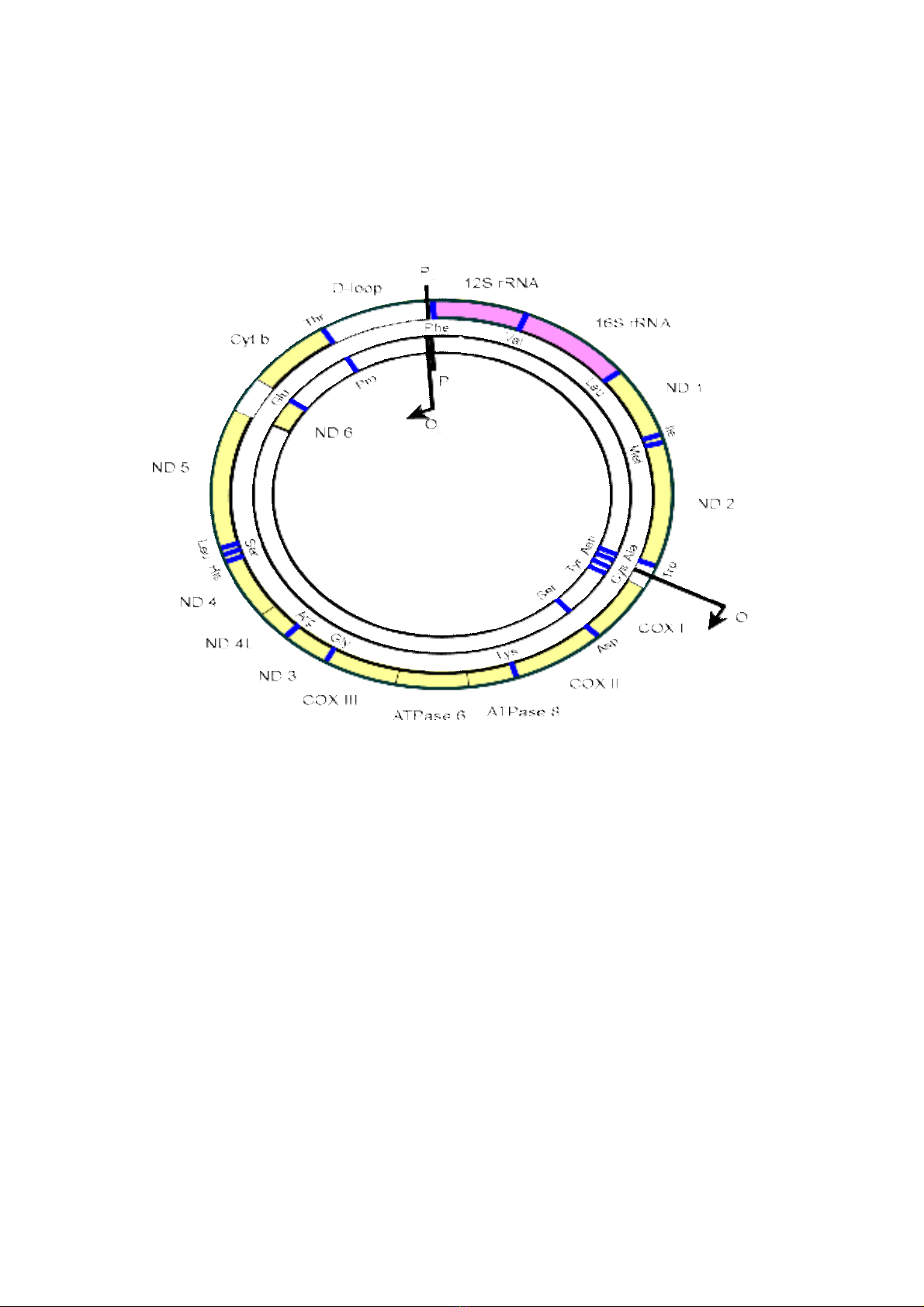
B gen ti th đc ký hi u là mtDNA (mitochondrial) d ng chu i xo n kép tr nộ ể ượ ệ ở ạ ỗ ắ ầ
(không có s tham gia c a các phân t histon), m ch vòng.Ti th có trong t bào t oự ủ ử ạ ể ế ạ
năng l ng cho t bào s d ng, mtDNA ch a trong nh ng túi nhi m s c th trongượ ế ử ụ ứ ữ ễ ắ ể
nhân c a ti th . mtDNA c a ti th ch a 37 gen, t t c mtDNA đóng vai trò thi t y uủ ể ủ ể ứ ấ ả ế ế
trong ch c năng c a ty th . mtDNA c a ti th có 13 gen tham gia vào t o enzym cóứ ủ ể ủ ể ạ
vai trò quan tr ng trong chu i chuy n đi n t phospholyl hóa. Quá trình này s d ngọ ỗ ể ệ ử ử ụ
oxygen đ thu 1 ph n năng l ng t các c u n i c a glucose đ gi năng l ng trongể ầ ượ ừ ầ ố ủ ể ữ ượ
ATP (andenosine triphosphate). ATP là ngu n năng l ng quan tr ng trong t bào.ồ ượ ọ ế
mtDNA còn l i trong ti th tham gia vào quá trình phiên mã t o ra RNA v n chuy n vàạ ể ạ ậ ể
RNA ribosome. Nh ng RNA này s tham gia vào quá trình d ch mã trong t bào ti thữ ẽ ị ế ể
t o ra protein và v n đm b o thông tin di truy n b gen c a ti th . [1]ạ ẫ ả ả ề ộ ủ ể
Kích th c mt DNA khác nhau tu loài[2]ướ ỳ
- N m men ấS. cerevisiae có mt DNA kích th c kho ng 84 kb ướ ả
- ng i, chu t và m t s đng v t h u nhũ kích th c mt DNA kho ng 16.5 kb.Ở ườ ộ ộ ố ộ ậ ữ ướ ả
- th c v t kích th c b gen ti th th ng r t l n ( ngô kho ng 570 kb) Ở ự ậ ướ ộ ể ườ ấ ớ ở ả
B gen ti th mtDNA mã hóa t ng h p cho nhi u thành ph n c a ti th : h th ng 2ộ ể ổ ợ ề ầ ủ ể ệ ố
l ai rRNA, 22-25 lo i tRNA và nhi u lo i protein có trong thành ph n c a màng bên tiọ ạ ề ạ ầ ủ
th . Trong khi đó, thành ph n l n protein c a ribosome ti th do các gen trong nhân xácể ầ ớ ủ ể
đnh ị
B gen ti th c a t bào đng v t g m các exon, b gen ti th cu t bào th c v tộ ể ủ ế ộ ậ ồ ộ ể ả ế ự ậ
và n m men g m các exon và intron xen k ấ ồ ẽ
B gen ti th đng v t h u nhũ có c u trúc t ng đi gi ng nhau, m i mtDNAộ ể ở ộ ậ ữ ấ ươ ố ố ỗ
g m 37 gen, trong đó có 13 gen mã hoá protein, 22 gen mã hoá tRNA và 2 gen mã hoáồ
rRNA.
Các gen ti th c a t bào đng v t h u nhũ phân b t ng đi không đng đu,ể ủ ế ộ ậ ữ ố ươ ố ồ ề
còn các gen ti th n m men t p trung m t c m 16 gen còn 10 gen phân tán kh p toànể ấ ậ ộ ụ ắ
b gen ti th .ộ ể
Ví d :ụ

* n m men DNA ti th có t l G-C kho ng 21%, còn DNA nhân có t l G-CỞ ấ ể ỷ ệ ả ỷ ệ
kho ng 40%.ả
* DNA ti th c a t bào đng v t có vùng gen bao trùm nhau (overlap), vùng D-loopể ủ ế ộ ậ
là vùng đi u khi n c a mtDNA. [2]ề ể ủ
Hình 6.1.1
B ng 6.1. M t s b ba mã hóa gen ti th các loài khác nhau.ả ộ ố ộ ể ở
Đa s các b nh di truy n đu do tác đng đn gene trong nhân. Tuy nhiên, có m tố ệ ề ề ộ ế ộ
s ít b nh là k t qu c a đt bi n trên ti th . ố ệ ế ả ủ ộ ế ể
B ba mã hóaộ
Mã di
truy nề
gen
nhân
Mã di truy n c a gen ti thề ủ ể
Đng v tộ ậ
h u nhũữ
Ru iồ
gi mấ
N mấ
men
Th c v tự ậ
UGA
AGA,AGG
AUA
AUU
CUU,CUC
Stop
Arg
Ile
Ile
Leu
Trp
Stop
Met
Met
Leu
Trp
Ser
Met
Met
Leu
Trp
Arg
Met
Met
Thr
Stop
Arg
Ile
Ile
Leu

M i t bào ng i th ng ch a hàng trăm ho c nhi u h n ti th trong t bào ch tỗ ế ườ ườ ứ ặ ề ơ ể ế ấ
c a chúng. Thông quá quá trình phosphoryl hóa oxi hóa ph c t p, nh ng bào quan nàyủ ứ ạ ữ
s n xu t ra ATP, ngu n năng l ng c n thi t cho s trao đi ch t t bào. Do đó ti thả ấ ồ ượ ầ ế ự ổ ấ ế ể
có vai trò quan tr ng cho s s ng t bào.ọ ự ố ế
Hình 6.1.2
* C u trúc gene ty th ng iấ ể ườ
B gen ti th ng i có kích th c kho ng 16.569bp, phân t m ch đôi, d ngộ ể ở ườ ướ ả ử ạ ạ
vòng đnh v trong matrix ti th và có kho ng vài nghìn b n sao trong m t t bào. DNAị ị ể ả ả ộ ế
ti th có 2 m ch, m ch n ng giàu guanine (H) và m ch nh giàu cytosine (L). M chể ạ ạ ặ ạ ẹ ạ
n ng ch a 12 trong 13 polypeptide mã hóa gene, 14 trong 22 gene tRNA và rRNA.ặ ứ
mtDNA không ch a intron và toàn b trình t đu mã hóa cho protein. Ch có m t vùngứ ộ ự ề ỉ ộ
không mã hóa trên mtDNA là D-loop, m t vùng 1121bp ch a đi m kh i đu sao chépộ ứ ể ở ầ
chu i H (OỗH) và promoter cho phiên mã chu i L và H. mtDNA đc sao chép t 2 đi mỗ ượ ừ ể
ori. S sao chép DNA kh i đu t Oự ở ầ ừ H s d ng m t RNA primer t o ra t s n ph mử ụ ộ ạ ừ ả ẩ
phiên mã chu i L. S t ng h p chu i H ti n hành kho ng 2/3 mtDNA, thay th chu iỗ ự ổ ợ ỗ ế ả ế ỗ
H b m cho đn khi nó ti n đn đi m ori chu i L (Oố ẹ ế ế ế ể ỗ L), trong c m kho ng 5 genụ ả
tRNA. Khi đng vào chu i H con, Oụ ỗ L cu n thành m t c u trúc thân-vòng và chu i Lộ ộ ấ ỗ
b t đu t ng h p.[2]ắ ầ ổ ợ
S phiên mã mtDNA kh i s t 2 promoter trong D-loop, Pự ở ự ừ L và PH. S phiên mãự
t c hai promoter t o ra m t RNA polycistronic. Gen tRNA xe gi a trình t rRNA vàừ ả ạ ộ ữ ự
mRNA sau đó s cu n l i và đc c t ra. mRNA và rRNA t do đc bi n đi sauẽ ộ ạ ượ ắ ự ượ ế ổ
d ch mã (g n đuôi polyA) và tRNA đc bi n đi đu 3’CCA.[2]ị ắ ượ ế ổ ầ
.
Hình 6.1.3
6.2. M i quan h ty th và nhân trong t bào. (ố ệ ể ế Nguy n Th Ph ng)ễ ị ươ
S l ng m ch polypeptide trong ti th m i loài r t khác nhau, m i loài cóố ượ ạ ể ở ỗ ấ ỗ
ch a h n 1 trăm m ch polypeptide. Khi dùng k thu t phân tích protein trong m iứ ơ ạ ỹ ậ ỗ
m ch polypeptide có ch a kho ng 500 – 800 proteins. Có ít nh t 100 gen trong nhânạ ứ ả ấ
góp ph n vào s t ng h p ch c năng ti th . Nhi u nguyên nhân ch a bi t rõ v m iầ ự ổ ợ ứ ở ể ề ư ế ề ố
quan h ti th và nhân nh ng có nhi u b ng ch ng xác đnh chúng có m i quan h v iệ ể ư ề ằ ứ ị ố ệ ớ
nhau trong quá trình s ng. Xác đnh protein trong ti th t nh ng trình t hoàn thànhố ị ể ừ ữ ự
nên h gen là n n t ng c b n tiên đón trình t đu tiên c quan bên trong t bào. Gi iệ ề ả ơ ả ự ầ ơ ế ớ
h n c a thành công này, là chu i n i liên ti p nhau b gián đo n trong gen m c tiêu vàạ ủ ỗ ố ế ị ạ ụ
t o ra nhi u protein có tín hi u nh n bi t r t th p. Nh ng nhóm gen và protein trongạ ề ệ ậ ế ấ ấ ữ
nhóm này có th chia thành 2 nhóm chính. Nhóm 1 gen c n thi t t ng h p s phát tri nể ầ ế ổ ợ ự ể

c a sinh v t bình th ng có đy đ ch c năng c a ty th . Nhóm 2 gen mã hóa proteinủ ậ ườ ầ ủ ứ ủ ể
đt bi n t o ra nh ng quy lu t khác nhau trong t ng h p sinh hóa h c đc bi t choộ ế ạ ữ ậ ổ ợ ọ ặ ệ
ho t đng c a ti th . M i quan h gen và ti th cung c p nhi u đi m nghi v n vạ ộ ủ ể ố ệ ể ấ ề ể ấ ề
nhi u d ng gen c a ti th và tr c đây ch a h bi t đn. [1]ề ạ ủ ể ướ ư ề ế ế
6.2.1. Nh ng enzym c n thi t cho s b o t n và bi u hi n c a h gen ti th .ữ ầ ế ự ả ồ ể ệ ủ ệ ể
Nhi u enzym c n thi t cho quá trình sao chép, phiên mã t o ra ti n RNA và d chề ầ ế ạ ề ị
mã t RNA tr ng thành. Các DNA polymerase thu c h enzym polymerase trong tiγừ ưở ộ ệ
th , ch có th t ng h p DNA b ng cách n i dài m t m i đã b t c p s n trên khuôn.ể ỉ ể ổ ợ ằ ố ộ ồ ắ ặ ẵ
M i này là m t RNA nh đc t ng h p b i m t ph c h p protein g i là primosome.ồ ộ ỏ ượ ổ ợ ở ộ ứ ợ ọ
Primosome bao g m nhi u protein và m t enzym t ng h p RNA t khuôn DNA g i làồ ề ộ ổ ợ ừ ọ
primase. Ngoài ra enzym này có ch c năng n i nh ng đo n ng n trên DNA đ t oứ ố ữ ạ ắ ể ạ
thành d ng m ch dài, d ng xo n, d ng vòng.[2]ạ ạ ạ ắ ạ
Enzym topoisomerases là enzym liên quan đn quá trình tách m ch và tái t h pế ạ ổ ợ
tích lũy c a DNA t o ra m i quan h quan tr ng cho nh ng ho t đng khác nhau củ ạ ố ệ ọ ữ ạ ộ ở ơ
quan c a sinh v t. Có nhi u ý ki n cho r ng trong ti th không có histone m c dùủ ậ ề ế ằ ể ặ
DNA c a ti th r t ph c t p và cũng đc d ch mã t o protein. DNA ty th t ngủ ể ấ ứ ạ ượ ị ạ ể ươ
đng v i vùng protein HU vi khu n và vùng protein HMG1 sinh v t nhân th t.ồ ớ ở ẩ ở ậ ậ
Trong d ch mã c n enzym RNA polymerase và ít nh t 1 đn 2 nhân t ho t hóa choị ầ ấ ế ố ạ
d ch mã. Nhi u lo i protein liên k t v i mtDNA t o ra c u trúc nucleod, m t s ho tị ề ạ ế ớ ạ ấ ộ ố ạ
đng liên k t v i RNA quá trình d ch mã đc kéo dài và d u hi u k t thúc m i loàiộ ế ớ ị ượ ấ ệ ế ở ỗ
sinh v t khác nhau. Vùng đc phiên mã g m các intron và exon; exon là trình t đcậ ượ ồ ự ượ
mã hóa c a gen và đc d ch mã thành protein. Intron là nh ng trình t đc phiên mãủ ượ ị ữ ự ượ
nh ng s b lo i b trong quá trong d ch mã. tRNAs, ribosome RNA cũng tham gia vàoư ẽ ị ạ ỏ ị
quá trình d ch mã RNA thành protein trong ti th . PolyA đc g n vào RNA d ch mãị ể ượ ắ ị
trong ti th c a đng v t đa bào thì đòi h i ít nh t 1 enzym g n vào.Trong t nhiên sể ủ ộ ậ ỏ ấ ắ ự ự
bi u hi n gen c a ti th c n 18 gen trong nhân, ngoài ra n m men trong quá trìnhể ệ ủ ể ầ ở ấ
v n chuy n đi n t c a cytochrome c c n thêm 1 gen n a. Gen này đc t o ra t quáậ ể ệ ử ủ ầ ữ ượ ạ ừ
trình đt bi n m t đo n trình t t ng h p ch c năng quan tr ng trong b gen ti thộ ế ấ ạ ở ự ổ ợ ứ ọ ộ ể
nh không có kh năng th c hi n quá trình hô h p c a t bào nh ng tham gia vàoư ả ự ệ ấ ủ ế ư
ch c năng quan tr ng nh t ng h p nhân heme, t ng h p lipid, chuy n hóa acid aminứ ọ ư ổ ợ ổ ợ ể
và t ng h p nhân t ch a Fe- S đóng vai trò co-factor trong m t s protein. S l ngổ ợ ố ứ ộ ố ố ượ
gen có ch c năng chính góp ph n t ng h p protein là enzym ti th ch y u ch aứ ầ ổ ợ ở ể ủ ế ứ

![Tài liệu học tập Chuyên đề tế bào [mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20250906/huutuan0/135x160/56151757299182.jpg)
![Câu hỏi ôn tập Sinh học tế bào [chuẩn nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20250709/kimphuong1001/135x160/771752031316.jpg)

![Lysosome là gì? - Nguyễn Huỳnh Thịnh [Giải thích A-Z]](https://cdn.tailieu.vn/images/document/thumbnail/2015/20151217/conheokiss/135x160/463746382.jpg)





















%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)