
Sinh học phân tử
134
Chương 7
Sửa chữa và bảo vệ DNA
Trên phân tử DNA có thể xuất hiện nhiều biến đổi do sai hỏng trong
quá trình trao đổi chất, do các tác nhân gây đột biến vật lý và hóa học của
môi trường. Tuy nhiên, genome luôn có độ ổn định cao nhờ các cơ chế sửa
chữa và bảo vệ DNA. DNA là phân tử duy nhất, mà khi biến đổi hay bị phá
hỏng vẫn có khả năng được sửa chữa nhờ tế bào. Các cơ chế sửa sai rất đa
dạng và có hiệu quả cao. Ba quá trình bao gồm sửa sai, tái bản và tái tổ hợp
DNA liên quan chặt chẽ với nhau. Đây cũng là một minh chứng về sự phối
hợp chặt chẽ giữa nhiều cơ chế di truyền.
I. Khái quát về các cơ chế sửa sai
Hầu hết các đột biến trên phân tử DNA thường được khắc phục bằng
hai phương thức chính:
- Sửa chữa phục hồi trực tiếp (direct reversal repair), hoặc:
- Cắt bỏ sai hỏng và sửa chữa lại bằng cách dùng trình tự bổ sung
(damage excision and repair using complementary sequence).
Sửa chữa trực tiếp thường liên quan đến hai loại sai hỏng trên phân tử
DNA do tia tử ngoại gây ra là: cyclobutane-pyrimidine dimer (CPDs) và
pyrimidine (6-4) pyrimidone (6-4 PPs). Hai loại sai hỏng này đều làm biến
dạng cấu trúc xoắn của DNA. CPDs và 6-4 PPs được nhận biết và sửa chữa
nhờ enzyme photolyase. Enzyme này sử dụng năng lượng ánh sáng để thực
hiện phản ứng làm thay đổi các liên kết hóa học để nucleotide trở lại dạng
bình thường. Phản ứng sửa chữa DNA bằng photolyase xảy ra trong rất
nhiều sinh vật prokaryote và eukaryote. Tuy nhiên, quá trình này không thấy
ở động vật có vú. Ở người cũng chưa phát hiện được bất kỳ loại photolyase
nào.
Đa số sai hỏng của DNA được sửa chữa bằng phương thức thứ hai. Cơ
chế này phải sử dụng thông tin di truyền chứa ở một trong hai sợi đơn DNA.
Khi trình tự nucleotide trên một sợi bị thay đổi thì sợi thứ hai (liên kết bổ

Sinh học phân tử
135
sung với sợi thứ nhất) được dùng làm khuôn mẫu để sửa chữa những sai
hỏng đó. Một số cơ chế sửa chữa như sau:
- Hệ thống sửa chữa nhận biết các trình tự DNA không thích hợp với
các cặp base chuẩn và thay thế chúng.
- Hệ thống sửa chữa-cắt bỏ (excision-repair system) loại đi một đoạn
DNA ở vị trí sai hỏng và sau đó thay thế nó.
- Hệ thống sửa chữa-tái tổ hợp (recombinant-repair system) sử dụng
phương thức tái tổ hợp để thay thế vùng sợi đôi bị sai hỏng.
Các hệ thống sửa chữa cũng phức tạp như bộ máy tái bản của nó, điều
đó cho thấy tầm quan trọng của chúng đối với sự sống của tế bào. Khi hệ
thống sửa chữa phục hồi một sai hỏng của DNA, thì không có một hậu quả
xấu nào xảy ra. Nhưng một đột biến có thể tạo ra hậu quả xấu khi DNA bị
hỏng.
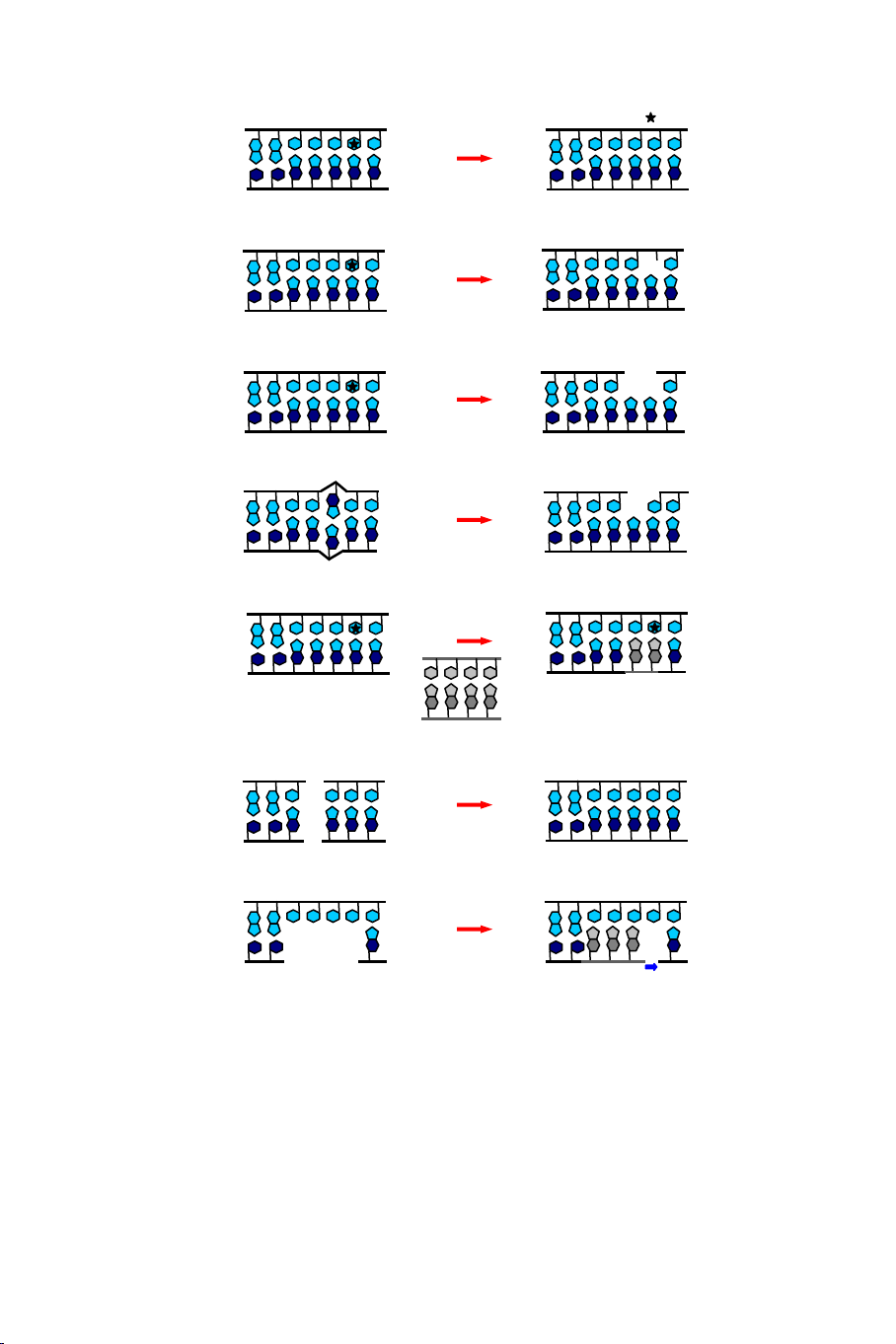
Hình 7.1 tóm tắt một số cơ chế sửa chữa DNA như sau:
- Một vài enzyme phục hồi trực tiếp các loại sai hỏng đặc biệt của
DNA.
- Một số phương thức sửa chữa bằng cách cắt bỏ base, sửa chữa bằng
cách cắt bỏ nucleotide, và sửa chữa ghép đôi lệch, tất cả chức năng của
chúng thực hiện bằng cách loại bỏ và thay thế nguyên liệu.
- Các hệ thống chức năng có thể tái tổ hợp để tạo ra một bản sao
không bị sai hỏng thay thế cho một sợi đôi bị sai hỏng.
- Phương thức nối đầu không tương đồng đã nối lại các đầu sợi đôi bị
hỏng.
- Một số DNA polymerase khác nhau cần thiết trong việc tổng hợp lại
các đoạn DNA thay thế.
1. Các biến đổi xảy ra trên phân tử DNA
Trên DNA có thể xảy ra các biến đổi ngẫu nhiên như sau:
- Gãy hay đứt mạch. Phân tử DNA có chiều ngang rất mảnh, bản thân
nó lại thường xuyên cuộn xoắn và giãn xoắn nên dễ xảy ra đứt gãy một sợi.
Tuy nhiên, khả năng đứt cùng lúc hai sợi hiếm khi gặp hơn.
Sinh học phân tử
136
Hình 7.1. Việc sửa chữa các gen có thể được phân loại theo các phương thức
sử dụng các cơ chế khác nhau để phục hồi hoặc bỏ qua sự sai hỏng của DNA
- Base bị cắt mất. Hiện tượng này làm base tương ứng không bắt cặp
được. Dưới tác dụng của nhiệt có thể xảy ra quá trình khử purine
(depurination) do thủy phân liên kết N-glycosyl.
Phục hồi trực tiếp sai hỏng
Sửa chữa bằng cách cắt bỏ base
Sửa chữa bằng cách cắt bỏ nucleotide
Sửa chữa bằng cách cắt bỏ ghép đôi lệch
Sửa chữa bằng cách tái tổ hợp
Nối đầu không tương đồng
DNA polymerase xúc tác các tiểu đơn vị

Sinh học phân tử
137
- Gắn các nhóm mới vào base bằng liên kết cộng hóa trị làm thay đổi
tính chất như trường hợp methyl hóa (gắn nhóm CH3 vào một base).
- Biến một base này thành một base khác làm bắt cặp sai. Ví dụ: quá
trình khử amine (desamination) của cytosine biến nó thành uracil.
- Các base có thể tồn tại ở hai dạng keto và enol nên có thể dẫn đến
bắt cặp sai. Ví dụ: dạng enol của cytosine có thể bắt cặp với adenine.
- Tạo các thymine dimer.
- Liên kết chéo giữa các mạch (interstand crosslink).
Trên đây là các biến đổi ngẫu nhiên, nếu có tác động của các tác nhân
gây đột biến các biến đổi sẽ xảy ra nhiều hơn.
2. Khái quát các cơ chế sửa chữa ở mức phân tử
Sự nguyên vẹn của phân tử DNA mang thông tin di truyền có ý nghĩa
sống còn đối với tế bào, nhất là tế bào prokaryote. Ở tế bào vi khuẩn, có đến
50% DNA không bị biến đổi thậm chí sau khi tái bản đến 100 triệu lần. Sự
ổn định cao của DNA tế bào có được nhờ hàng loạt các cơ chế bảo vệ sự
nguyên vẹn và sửa chữa ngay lập tức bất kỳ sai hỏng nào vừa xuất hiện.
Các hệ thống sửa sai rất đa dạng và có hiệu quả rất cao ở E. coli. Có
khoảng 100 locus tham gia trực tiếp hoặc gián tiếp vào việc bảo vệ DNA và
sửa sai.
Chỉ với DNA, tế bào phải đầu tư rất lớn cho sự ổn định của thông tin
di truyền. Nếu như sự tái bản chỉ xảy ra khi phân bào, thì các cơ chế sửa sai
phải hoạt động liên tục. Tái bản được thực hiện với cơ chế về cơ bản giống
nhau ở các loại tế bào và trong mỗi lần nhân đôi DNA. Trong khi đó, sai
hỏng xảy ra rất đa dạng, nên các cơ chế sửa sai nhiều hơn và với số lượng
gen tham gia lớn hơn. Hơn nữa, các cơ chế di truyền cơ bản như tái bản, tái
tổ hợp DNA đều có sự tham gia của các cơ chế sửa sai và ngược lại sửa sai
cần đến tái bản và tái tổ hợp.
3. Biến đổi làm tăng tần số đột biến
Nhờ các cơ chế sửa sai nên bình thường DNA tế bào có độ ổn định rất
lớn, thường sự thay thế base có tần số 10-9-10-10. Tuy nhiên, người ta đã phát
hiện được các dòng đột biến ngẫu nhiên tăng cao hơn hẳn, chúng được gọi

Sinh học phân tử
138
là mutator (nhân tố gây đột biến). Trong nhiều trường hợp, các kiểu hình
mutator liên quan đến sai hỏng trong hệ thống sửa sai. Ở E. coli, các locus
mutator mutH, mutL, mutU và mutS tác động đến các cấu phần của hệ thống
sửa sai do ghép đôi lệch (mismatch-repair) sau tái bản nên đã làm cho tần số
đột biến ngẫu nhiên tăng cao.
Ngoài mutator, sự sai hỏng làm tăng tính nhạy cảm với tia tử ngoại
(ultraviolet) và tăng sai hỏng khi tái tổ hợp.
II. Các kiểu sửa chữa
1. Quang tái hoạt hóa
Sửa chữa trực tiếp bằng quang tái hoạt hóa (photoreactivation) ít khi
gặp, nó bao gồm sự phục hồi hoặc loại bỏ đơn giản các sai hỏng và quá trình
này xảy ra ở ngoài sáng. Quang tái hoạt hóa của các pyrimidine dimers,
trong đó tạo cơ hội cho các liên kết cộng hóa trị được phục hồi nhờ enzyme
phụ thuộc ánh sáng (light-dependent enzyme), là một ví dụ điển hình (Hình
7.2). Hệ thống sửa chữa này phổ biến trong tự nhiên, và đặc biệt quan trọng
ở thực vật. Trong E. coli, nó phụ thuộc vào sản phẩm của một gen đơn (phr)
mã hóa cho một enzyme được gọi là photolyase.
Sau khi xử lý tia tử ngoại gây đột biến, nếu đưa ra ánh sáng thì phần
lớn sai hỏng được phục hồi. Hiện tượng này được gọi là quang tái hoạt hóa.
Năng lượng của ánh sáng khả kiến (từ 300 đến 600 nm) hoạt hóa photolyase
(gen phr ở E. coli) cắt các vòng cyclobutyl pyrimidine dimer (thường là
thymine dimer).
Enzyme này hoạt động trong các tế bào ở nhiều loài khác nhau. Vào
ban ngày, các sinh vật thường chịu tác động của ánh sáng, nên cơ chế quang
tái hoạt hóa (quang phục hồi) có vai trò quan trọng trong sửa sai DNA. Ví
dụ: Mycoplasma, sinh vật đơn giản nhất hiện nay, chỉ có vài trăm gen nhưng
một trong số đó đã được dùng cho quang tái hoạt hóa.
Ngoại trừ sửa chữa bằng quang tái hoạt hóa xảy ra ngoài sáng, phần
lớn các cơ chế sửa chữa khác được thực hiện trong tối, nhờ các enzyme
nuclease cắt bỏ chỗ sai hỏng theo nhiều cách như sau:
2. Sửa chữa ghép đôi lệch
Sửa chữa ghép đôi lệch (mismatch-repair) giữa các sợi DNA là một
trong những mục tiêu chính của hệ thống sửa sai. Sửa chữa ghép đôi lệch


![Bài giảng Kỹ thuật DNA và công nghệ sinh học [mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2022/20220110/trollhunters/135x160/9101641828200.jpg)























%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)