
Effects of a tryptophanyl substitution on the structure and
antimicrobial activity of C-terminally truncated gaegurin 4
Hyung-Sik Won
1
, Sang-Ho Park
1
, Hyung Eun Kim
1
, Byongkuk Hyun
2
, Mijin Kim
2
, Byeong Jae Lee
2
and
Bong-Jin Lee
1
1
College of Pharmacy, Seoul National University, Seoul, South Korea;
2
Institute of Molecular Biology and Genetics,
Seoul National University, Seoul, South Korea
Gaegurin 4 (GGN4), a 37-residue antimicrobial peptide,
consists of two amphipathic ahelices (residues 2–10 and
16–32) connected by a flexible loop region (residues 11–
15). As part of an effort to develop new peptide antibiotics
with low molecular mass, the activities of C-terminally
truncated GGN4 analogues were tested. D
24)37
GGN4, a
peptide analogue with 14 residues truncated from the
C-terminus of GGN4, showed a complete loss of anti-
microbial activity. However, the single substitution of
aspartic acid 16 by tryptophan (D16W) in the D
24)37
GGN4 completely restored the antimicrobial activity,
without any significant hemolytic activity. In contrast,
neither the D16F nor K15W substitution of the D
24)37
GGN4 allowed such a dramatic recovery of activity. In
addition, the D16W substitution of the native GGN4
significantly enhanced the hemolytic activity as well as the
antimicrobial activity. The structural effect of the D16W
substitution in the D
24)37
GGN4 was investigated by CD,
NMR, and fluorescence spectroscopy. The results showed
that the single tryptophanyl substitution at position 16 of
the D
24)37
GGN4 induced an ahelical conformation in the
previously flexible loop region in intact GGN4, thereby
forming an entirely amphipathic ahelix. In addition, the
substituted tryptophan itself plays an important role in the
membrane-interaction of the peptide.
Keywords: antimicrobial peptide; GGN4 analogues; try-
ptophanyl substitution; CD; NMR.
Membrane-active peptides exhibit many interesting biolo-
gical and pharmacological activities, and they can also serve
as model systems for large membrane proteins [1]. Partic-
ularly, many organisms, including fungi, insects, amphibi-
ans, and humans, produce hydrophobic and amphipathic
peptides that exhibit antibiotic, fungicidal, hemolytic, viru-
cidal, and tumoricidal activities. Now, it is becoming clear
through many studies that the antimicrobial peptides are an
important component of the innate defenses of all species of
life [2–8]. Presently, more than 100 molecules with this
property have been isolated from various vertebrates as well
as invertebrates. These antimicrobial peptides can be
grouped into three classes, depending on their structural
properties [9]: ahelicoidal peptides, peptides with one to
several disulfide bridges, and peptides rich in certain amino
acids such as Proline or Tryptophan. Most of these peptides
share some common characteristics, such as their low
molecular mass (2–5 kDa), the presence of multiple lysine
and arginine residues, and their amphipathic nature.
Although the exact mechanism by which they kill bacteria is
not clearly understood, it has been shown that peptide–lipid
interactions leading to membrane permeation play a role in
their activity.
The best understood group includes the linear amphi-
pathic ahelical antimicrobial peptides [1,10–13]. Although
most of these peptides dissolve well in aqueous solutions,
they also show a strong affinity for phospholipid mem-
branes. Generally, they adopt a highly ordered helical
structure in hydrophobic or membrane-mimetic environ-
ments, whereas they assume a random coil conformation in
aqueous solutions. It has been demonstrated that the
structural and physico-chemical properties, such as the
amino-acid composition, helical length, and amphipathic
nature, etc. of the peptides, rather than the primary
sequence similarity or specific receptor–ligand interactions,
are responsible for their biological activity [1]. Two plausible
models for the membrane permeation mechanism by
amphipathic ahelical peptides have been proposed [10]:
the barrel-stavemechanism¢and the carpet-likemechan-
ism. In the former, the transmembrane amphipathic aheli-
ces form bundles, producing a transmembrane pore. The
latter describes membrane disintegration by disruption of
the bilayer curvature, leading to micellization. In this model,
in contrast to the barrel-stave mechanism, the peptides do
not penetrate into the hydrophobic core of the membrane,
but rather bind to the phospholipid headgroups.
A number of peptides with a broad-spectrum of antimi-
crobial activities have been isolated from the skin of various
amphibians, and six antimicrobial peptides, named gaegu-
rins (GGNs), were also isolated from the skin of a Korean
frog, Rana rugosa [14]. Some of them, particularly those
with no or little hemolytic activity, are considered as target
molecules for the development of new antibiotic or
Correspondence to B.-J. Lee, College of Pharmacy, Seoul National
University, San 56-1, Shillim-Dong, Kwanak-Gu, Seoul 151-742,
South Korea. Fax: + 82 2872 3632, Tel.: + 82 2880 7869,
E-mail: lbj@nmr.snu.ac.kr
Abbreviations: DPC, dodecylphosphocholine; GGN4, gaegurin 4;
MIC, minimal inhibitory concentration; NATA, N-acetyl-
L
-tryp-
tophanamide; TFE, 2,2,2-trifluoroethanol; [q]
M
, mean residue molar
ellipticity.
(Received 3 June 2002, accepted 25 July 2002)
Eur. J. Biochem. 269, 4367–4374 (2002) FEBS 2002 doi:10.1046/j.1432-1033.2002.03139.x

anticancer agents by peptide engineering. Out of the six
gaegurins, GGN4 has the longest length and is the most
abundant in the frog skin. Thus, the peptide is believed to be
crucial in the innate defense system of the frog. Our previous
work [15] showed that GGN4 adopts a random structure in
an aqueous solution, but adopts a helical conformation
consisting of two amphipathic ahelices (residues 2–10 and
16–32) in membrane-mimetic environments. Recently, as
part of an effort to develop new potential peptide antibiotics
with lower molecular mass, the antimicrobial activities of
several GGN4 analogues with C-terminal truncations were
analyzed [16]. The deletion of up to 14 residues from the
C-terminus of GGN4 almost completely abolished the
antimicrobial activity of the peptide, but the concomitant
single substitution of aspartic acid 16 with tryptophan
showed a nearly complete restoration of activity.
In the present work, we further examined the biological
activities of several GGN4 analogue peptides. The struc-
tural effect and the functional role of the tryptophanyl
substitution at position 16 was investigated for the
C-terminally truncated GGN4, by CD, fluorescence, and
nuclear magnetic resonance (NMR) spectroscopy. We
expect that the present results will not only improve our
understanding of the action mechanism of antimicrobial
peptides, but also present new perspectives for the develop-
ment of new peptide antibiotics.
EXPERIMENTAL PROCEDURES
Materials, peptide preparation, and activity test
N-Acetyl-
L
-tryptophanamide (NATA), 2,2,2-trifluoroetha-
nol-d
3
99.5% (TFE-d
3
), and sodium dodecyl-d
25
sulfate
(SDS-d
25
) were obtained from Aldrich. D
2
O (99.95%) was
obtained from Sigma, and all other chemicals were either
analytical or biotechnological grade. GGN4 analogue
peptides were purchased from ANYGEN (Kwang-ju,
Korea; URL, http://www.anygen.com). The sequence and
purity of the peptides were confirmed by mass spectrometry
and high performance liquid chromatography. Antimicro-
bial activities of the peptides were determined by measuring
the minimal inhibitory concentrations (MIC) for diverse
microorganisms, as described previously [14,15]. Hemolytic
activities of the peptides were estimated as the percent
hemolysis relative to that by 0.1% Triton X-100, as
described by Park et al. [14].
CD and fluorescence spectroscopy
For CD spectroscopy, the peptide powder was dissolved to
a final concentration of 50 l
M
, in various solvents: 20 m
M
sodium acetate buffer (pH 4.0), TFE/water mixtures, 5 m
M
DPC micelles, and 10 m
M
SDS micelles. Before the CD
measurement, the pH was adjusted to 4.0 by the addition of
0.1
M
HCl or NaOH. CD spectra were obtained at 20 Con
a JASCO J-720 spectropolarimeter, using a 0.2-cm path-
length cell, with a 1-nm bandwidth and a 4-s response time.
CD scans were taken from 250 nm to 190 nm, with a scan
speed of 50 nmÆmin
)1
and a 0.5-nm step resolution. Three
scans were added and averaged, followed by subtraction of
the CD signal of the solvent. Finally, the CD intensity was
normalized by the equation as the mean residue molar
ellipticity:
½hk
M¼hk105
lcn
where ½hk
M(deg cm
2
Ædmol
)1
)andhk(mdeg) are the mean
residue molar elipticity and the observed CD intensity at
any wavelength (k), respectively. l,c,andnrepresent the
path-length (cm), the concentration (l
M
), and the number
of residues, respectively.
Fluorescence emission was monitored on a Hitachi
F-4500 fluorimeter, between 300 and 450 nm at 0.2 nm
increments, with an excitation wavelength of 280 nm, using
a 10-mm quartz cell at room temperature. Scans were taken
with a 5-nm excitation and emission bandwidth, a 0.5-s
response time, and a scan speed of 40 nmÆs
)1
. All samples
contained 8 l
M
peptide or the same concentration of
NATA for control experiments, in water or a 10-m
M
SDS
solution at pH 4.0. All spectra were baseline corrected by
subtracting the corresponding solvent spectrum.
NMR Spectroscopy and structure calculation
Samples for NMR measurements contained 5 m
M
peptide
in TFE-d
3
/H
2
O (1 : 1, v/v) at pH 4.0, and in 500 m
M
SDS-
d
25
at pH 4.0. NMR spectra were recorded on a Bruker
DRX-500 spectrometer, at 298 K in 50% TFE/water and at
313 K in SDS micelles. Solvent suppression was achieved
using selective low-power irradiation of the water resonance.
The 2D TOCSY spectra were acquired with an isotropic
mixing time of 60 ms. The 2D NOESY spectra were
acquired with mixing times of 150 and 200 ms, respectively.
Slowly exchanging amide protons were monitored by the
D
2
O exchange experiments with a series of 2D NOESY
spectra measured immediately after the addition of deuter-
ated solvent to a sample lyophilized from nondeuterated
solvent, as described previously [15,17]. In order to study the
interaction between the peptide and SDS micelles, the 2D
NOESY spectrum was acquired at 313 K, for 2.5 m
M
of the
peptide dissolved in a solution containing 20 m
M
nondeu-
terated SDS micelles at pH 4.0, with a 200-ms mixing time.
The suppression of the water signal was achieved by the
pulsed field gradient method. All NMR spectra were
processed and analyzed using the
NMRPIPE
/
NMRDRAW
software and the
NMRVIEW
program [18,19]. Sequence-
specific assignments of the proton resonances were achieved
by spin system identification from the TOCSY and DQF-
COSY spectra, followed by sequential assignments through
the NOE connectivities [15,17,20]. Distance restraints,
backbone dihedral angle restraints, and hydrogen bond
restraints were obtained and used for the structure calcu-
lation by the simulated annealing and energy minimization
protocol in the program
XPLOR
3.851 [21], as described
previously [15]. Out of the 50 structures calculated by the
method demonstrated previously [15], the 49 accepted
structures were refined, and finally 20 structures with the
lowest energies were chosen to represent the solution
structure.
RESULTS AND DISCUSSION
Biological activities of the GGN4 analogues
Native GGN4 exhibits a broad range of antimicrobial
activity against prokaryotic cells, but very little hemolytic
4368 H.-S. Won et al. (Eur. J. Biochem. 269)FEBS 2002

activity against human red blood cells [14,15]. As shown in
Table 1, the C-terminal 14 residue truncated GGN4 (D
24)37
GGN4) showed neither antimicrobial activity against
bacterial cells nor hemolytic activity against human red
blood cells. Surprisingly, D16W-D
24)37
GGN4, a GGN4
analogue with both the C-terminal 14 residue truncation
and the substitution of the aspartic acid at position 16 by
tryptophan, showed antimicrobial activity comparable to
that of native GGN4 and less hemolytic activity than that of
native GGN4. These results are consistent with the previous
report by Kim et al. [16], in which the antimicrobial
activities were checked against only two species of bacteria
(Micrococcus luteus and Escherichia coli). In this previous
report, the antimicrobial activities of several C-terminally
truncated GGN4 analogues with a substituted tryptophan
were analyzed. The single tryptophanyl substitution of the
C-terminally truncated GGN4, at position 3, 17, 18, or 19,
did not increase the activity. Likewise, in the present work,
the tryptophanyl substitution at position 15 (K15W-D
24)37
GGN4) did not restore the antimicrobial activity. Taken
together, these results suggest that position 16 is the most
effective position for a single tryptophanyl substitution to
increase the antimicrobial activity of the C-terminally
truncated GGN4. In addition, in this work, the single
phenylalanine substitution at position 16 of the C-terminally
truncated GGN4 moderately restored the activity of the
peptide, but less than that by tryptophan. This suggests that
the single tryptophan introduced at position 16 of the
D16W-D
24)37
GGN4 would have an amino-acid specific
role in the biological action of the peptide. Finally, the effect
of the tryptophanyl substitution at position 16 on the
biological activity was confirmed for the native GGN4.
Consistent with the results of the C-terminally truncated
GGN4, the D16W substitution in the native GGN4 also
significantly increased the antimicrobial activity of the
peptide. However, a remarkable increase of the hemolytic
activity was observed concomitantly.
Conformational preferences of the GGN4 analogues
Figure 1 summarizes the CD results of the GGN4 analogue
peptides in aqueous buffer and membrane-mimetic envi-
ronments (50% TFE/water, 10 m
M
SDS micelles, and
5m
M
DPC micelles). For clarity, ½h222
M
jj
, the absolute value
of the mean residue molar elipticity at 222 nm, which
approximately reflects the helical content [13,15,17], is
indicatedintheinsetofeachpanel.½h208
M=½h222
M,theratioof
mean residue molar elipticity at 208 nm (½h208
M)tothatat
222 nm (½h222
M), is also included in parentheses, in order to
reflect the spectral shape. In aqueous buffer, the CD spectra
of the GGN4 analogues, including the native GGN4,
showed a strong negative band near 200 nm and a weak and
broad band around 222 nm, indicating a predominantly
random-coil conformation with a slight helical propensity
[17,22,23]. Especially, the D16W GGN4 showed a rather
significant helical content, even in the aqueous buffer.
However, in a 50% TFE/water mixture, the CD spectra
changed dramatically, with a strong positive band near
192 nm and strong negative bands centered at 208 and
222 nm, which are indicative of a highly ahelical confor-
mation [22–25]. The signals at 193, 208, and 222 nm were
intensified with increasing percentages of TFE, which
indicates that the helicity of the peptides increased within
more hydrophobic environments. The spectral change
induced by the increased concentration of TFE was nearly
complete at about 40–60% TFE/water, and no significant
spectral change occurred upon the change of pH from 3.0 to
7.0 in the 50% TFE/water solution (data not shown). The
CD spectra in 10 m
M
SDS and 5 m
M
DPC micelles, which
are above their critical micellar concentrations [12,26,27],
showed shapes similar to those in 50% TFE/water, also
indicating a typical ahelix pattern. This conformational
change from a random-coil in aqueous buffer to an ahelix
in membrane-mimetic environments is common to many
membrane-binding peptides [1,10–13,17].
Although all of the GGN4 analogues tested in this work
showed the same conformational preferences in various
solvents, they differed remarkably from one another in
their helical contents deduced from ½h222
M
jj
andinthedetailed
spectral shape represented by ½h208
M=½h222
M.Thesetwoparam-
eters, ½h222
Mand ½h208
M=½h222
M, correlated well with the biological
activities. In membrane mimetic environments (50% TFE,
10 m
M
SDS, and 5 m
M
DPC), among the C-terminally
truncated GGN4 analogues, D16W-D
24)37
GGN4, which
exhibited the largest antimicrobial activity, showed the
largest ½h222
M
jj
and a relatively small ½h208
M=½h222
M, while D
24)37
Table 1. Antimicrobial activity (a) and hemolytic activity (b) of GGN4 analogue peptides. Percent hemolysis is relative to that by 0.1% Triton X-100.
Molecular masses (in Da) are: Native, 3748; D16W, 3819; N
23
, 2358; D16W-N
23
, 2429; D16F-N
23
,2390;K15W-N
23
, 2416.
Microorganism Native D16W N
23
D16W-N
23
D16F-N
23
K15W-N
23
Minimal inhibitory concentration values (lgÆmL
)1
):
Micrococcus luteus 2.5 25 > 200 2.5 > 200 > 200
Bacillus subtilis 10 2.5 > 200 25 25 > 200
Klebsiella pneumoniae 25 10 > 200 25 100 > 200
Shigella dysentariae 25 10 > 200 50 50 > 200
Pseudomonas aeruginosa 100 50 > 200 125 200 > 200
Escherichia coli 75 10 > 200 25 25 > 200
Salmonella typhimurium 200 50 > 200 125 > 200 > 200
Serratia marcescens >200 >200 >200 >200 >200 >200
Percent hemolysis values:
10 lgÆmL
)1
concentration 0.78% 1.97% 0% 0.02% 0.06% 0%
100 lgÆmL
)1
concentration 1.67% 52.9% 0% 0.38% 0.32% 0%
FEBS 2002 Structure–activity relationships of GGN4 analogues (Eur. J. Biochem. 269) 4369
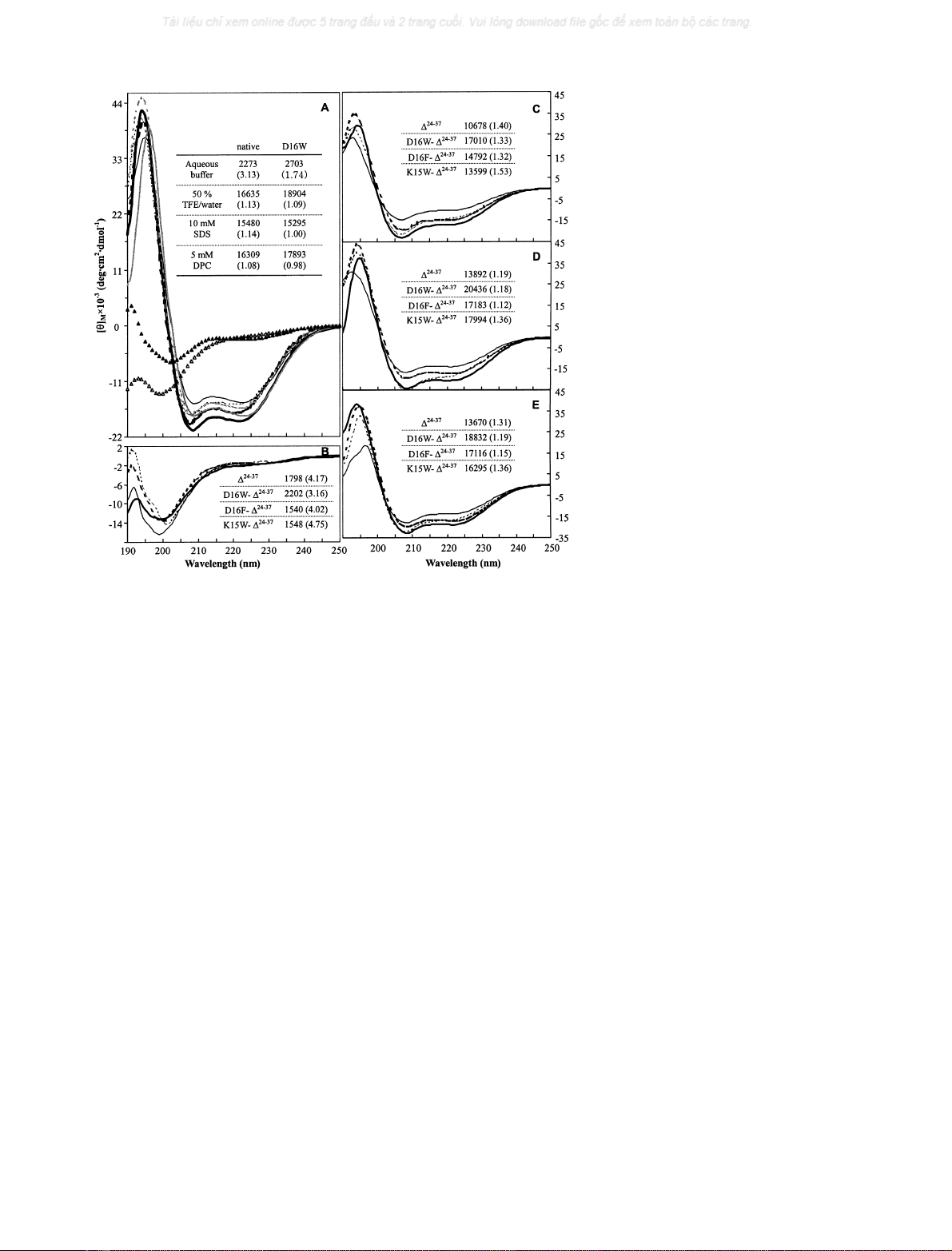
GGN4, which exhibited no significant activity, showed the
least ½h222
M
jj
and a relatively large ½h208
M=½h222
M.The ½h222
M
jj
of
D16F-D
24)37
GGN4, which showed moderate activity, was
between that of D
24)37
GGN4 and that of D16W-D
24)37
GGN4. The ½h208
M=½h222
Mof D16F-D
24)37
GGN4 was also
relatively small. In contrast, K15W-D
24)37
GGN4, which
has no activity, showed the largest ½h208
M=½h222
Mand a relatively
small ½h222
M
jj
.
Generally, SDS micelles, which have negatively charged
surfaces, mimic the bacterial cell membrane with its
negatively charged surface, while DPC micelles, which
have zwitterionic surfaces, mimic the eukaryotic cell
membrane with its zwitterionic surface [10,17,28]. In the
case of the C-terminally truncated GGN4 analogues, the
maximum ½h222
M
jj
and the minimum ½h208
M=½h222
Mwere com-
monly observed in SDS micelles. However, both the
native GGN4 and D16W GGN4 showed a higher ½h222
M
jj
and a lower ½h208
M=½h222
M
jj
in DPC micelles than those in SDS
micelles. In particular, in DPC micelles, D16W GGN4,
which was the only peptide with significant hemolytic
activity as well as the largest antimicrobial activity, yielded
a½h208
M=½h222
Meven lower than that of native GGN4, as well
as a larger ½h222
M
jj
than that of native GGN4.
In summary, the CD results showed that the differences
in the activities between the GGN4 analogue peptides are
deeply related to their conformational properties and helical
contents in various environments. In addition, it became
clear that the D16W substitution of both the native and the
C-terminally truncated GGN4 increased the helical
propensity of the peptides, which would have a key role in
increasing their biological activities.
Solution structures of GGN4 analogues
In order to reveal the detailed structural effects of the D16W
substitution, the solution structures of D
24)37
GGN4 and
D16W-D
24)37
GGN4 were investigated by NMR spectros-
copy. The structure of the native GGN4 in 50% TFE/water
consists of two ahelices extending from residues I2 to A10
and from residues D16–32, respectively [15]. The final
selected structures (Fig. 2A) and the refined average struc-
ture (figure not shown) of D
24)37
GGN4 in 50% TFE/water
reveal the well-ordered N-terminal ahelix composed of
residues from I2 to K11, which is in good agreement with
the corresponding part of the native GGN4. However, the
C-terminal part (residues 12–23) of D
24)37
GGN4 showed
no significant secondary structure, although some of the
initially solved 50 structures randomly showed a short
ahelical conformation in the C-terminal part. The dis-
ordered conformation in the C-terminal region of D
24)37
GGN4 is probably due to the break of the peptide bond at
position 23. In contrast, the finally selected structures
(Fig. 2B) and the refined average structure (Fig. 3A) of
D16W-D
24)37
GGN4 showed a stable helical conformation
from residues I2 to V18 in 50% TFE/water, although a few
of the initially solved 50 structures randomly showed a
rather loosened conformation in the C-terminal part. In the
previous work [15], the loop region (residues 11–15) of
native GGN4 exhibited a flexible, but helix-like conforma-
tion in the membrane-mimetic environment, although it
could not be defined as a stable a-helix. However, the
corresponding region in D16W-D
24)37
GGN4 showed a
stable ahelical conformation joined to its N- and C-terminal
Fig. 1. Conformational preferences of GGN4
analogues in various solvents. A: CD spectra of
native (empty symbols and broken lines) and
D16W GGN4 (filled symbols and solid lines)
in aqueous buffer (triangle symbols), 50%
TFE/water mixture (bold lines), 10 m
M
SDS
micelles (gray lines), and 5 m
M
DPC micelles
(thinlines).B–E:CDspectraofD
24)37
(thin,
solid line), D16W-D
24)37
(bold, solid line),
D16F-D
24)37
(bold, broken line), and K15W-
D
24)37
GGN4 (thin, broken line), in aqueous
buffer (B), 50% TFE/water mixture (C),
10 m
M
SDS micelles (D), and 5 m
M
DPC
micelles(E).Ineachpanel, ½h222
M
jj
and ½h208
M=½h222
M
(in parentheses) of each sample are tabulated
in the inset.
4370 H.-S. Won et al. (Eur. J. Biochem. 269)FEBS 2002

portions. Position 16 is at the border between the loop
region (residues 11–15) and the C-terminal helix (residues
16–32) in the intact GGN4. Thus, it can be inferred that the
W16 residue of D16W-D
24)37
GGN4 would stabilize the
potential helical propensity of the previous loop region as
well as the C-terminal helix destabilized by truncation. In
line with the CD results, the solution structures clearly
support the idea that the D16W substitution of D
24)37
GGN4 contributed to the restoration of the antimicrobial
activity, at least by changing its structure.
In order to elucidate the structure-function relationship
of D16W-D
24)37
GGN4, its structure in SDS micelles was
also investigated. The helical structures of D16W-D
24)37
GGN4 in SDS micelles and in TFE/water were very similar
to each other (Fig. 2), and showed several structural
features that are characteristic of many membrane-binding
peptides. To begin with, as shown in Fig. 3, the peptide
adopts a typical amphipathic helix structure, with the
hydrophobic residues on one side and the hydrophilic
residues on the other side of the helical axis. In particular, all
of the lysine residues are oriented to the same side. Thus, it
can be deduced that the positively charged hydrophilic side
would easily recognize and bind to the negatively charged
membrane surface of microorganisms. Indeed, in the
NOESY experiment of D16W-D
24)37
GGN4 in SDS
micelles, we observed intraresidue NOEs between the side-
chain H
e
and Hz atoms of lysine residues (data not shown),
which could not observed in the TFE/water mixture,
probably due to the high mobility of the side-chain or the
rapid exchange of the Hz amino protons. This observation
indicates that the lysine side-chains are immobilized in SDS
micelles, probably by the electrostatic interaction between
their positively charged amino groups and the negatively
charged surfaces of the SDS micelles. In addition, consistent
with the CD results, D16W-D
24)37
GGN4 displayed a more
lengthened a-helix (from I2 to G20) in SDS micelles than
that in 50% TFE/water (Fig. 2). The relatively more stable
C-terminal helical structure of D16W-D
24)37
GGN4 in SDS
micelles than in TFE/water is also attributable to the
possible interaction between the K19 residue and the SDS
micelles.
In many cases, the amphipathic nature of a helical
peptide is known to be important for its membrane binding
[1,10]. Along with the positively charged side-chains from
the hydrophilic face, the nonpolar residues in the hydro-
phobic face of D16W-D
24)37
GGN4 seem to contact the
SDS micelles by hydrophobic interactions with the acyl
chains of the micelles. This possible interaction, which has
been proposed for other amphipathic peptides [1,10,17,24],
is also supported in this work by the intermolecular NOEs
between several hydrophobic residues of the peptide and the
acyl chains of the SDS molecules. In the NOESY experi-
ment of D16W-D
24)37
GGN4 in the nondeuterated SDS
micelles (Fig. 4), a strong resonance at about 1.19 p.p.m.,
which originates from the methylene protons of SDS [29],
was observed. Figure 4 clearly depicts the intermolecular
NOE cross-peaks between the SDS methylene protons and
the peptide backbone amide protons of the F9, V13, and
Fig. 2. Solution structures of GGN4 analogues. Backbone atoms (N, C
a
,andC¢) of the finally refined 20 structures were superimposed, by matching
the backbone atoms in the helical region, for D
24)37
GGN4 in the 50% TFE/water mixture (A), D16W-D
24)37
GGN4 in the 50% TFE/water
mixture (B), and D16W-D
24)37
GGN4 in SDS micelles (C), respectively. In panel D, the set of D16W-D
24)37
GGN4 structures in the 50% TFE/
water mixture (gray lines) was superimposed over that in 500 m
M
SDS micelles (black lines), by matching the backbone atoms in residues I2V18,
andthemainchain(N,C
a
,C¢, and O) and the tryptophan side chain atoms are represented.
Fig. 3. Refined average structure of D16W-
D
24)37
GGN4. Residues 2–19 in the 50% TFE/
water mixture (A and C) and residues 2–20 in
500 m
M
SDS micelles (B and D) are shown as
space-filling models. Hydrophilic, hydropho-
bic, and tryptophan residues are colored
black, gray, and dark gray, respectively. The
direction of view is approximately perpen-
diculartothehelicalaxisinpanelsAandB,
and is parallel to the helical axis in panels C
and D.
FEBS 2002 Structure–activity relationships of GGN4 analogues (Eur. J. Biochem. 269) 4371


























%20--%3e%3cdefs%3e%3cstyle%3e%20.st0%20{%20fill:%20%23fff;%20}%20.st1%20{%20fill:%20%237800fa;%20}%20%3c/style%3e%3c/defs%3e%3cpath%20class='st1'%20d='M117.78,12.18H43.11c2.9,3.47,4.65,7.94,4.65,12.82,0,5.6-2.3,10.66-6.01,14.29h76.02l7.22-13.56-7.22-13.56Z'/%3e%3cg%3e%3cpath%20class='st0'%20d='M53.58,26.17h-.59v-1.46h.59v-4.96h2.83c1.78,0,2.67.94,2.67,2.82v5.76c0,1.87-.89,2.81-2.67,2.81h-2.83v-4.96ZM55.36,21.37v3.34h1.1v1.46h-1.1v3.34h1.01c.61,0,.91-.37.91-1.1v-5.93c0-.74-.3-1.1-.91-1.1h-1.01Z'/%3e%3cpath%20class='st0'%20d='M65.99,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM65.28,18.04c-.25.46-.51.77-.75.94-.21.15-.47.22-.79.22-.26,0-.57-.07-.92-.22l-.38-.15c-.14-.05-.26-.07-.37-.07-.3,0-.53.18-.71.54l-.91-.68c.25-.46.51-.77.75-.94.21-.14.48-.21.79-.21.26,0,.57.07.92.21l.38.15c.14.05.26.07.37.07.3,0,.53-.18.71-.54l.91.68ZM61.91,27.52h1.73l-.87-5.76-.87,5.76Z'/%3e%3cpath%20class='st0'%20d='M74.53,26.89v1.52c0,1.91-.89,2.86-2.67,2.86s-2.67-.95-2.67-2.86v-5.93c0-1.91.89-2.86,2.67-2.86s2.67.95,2.67,2.86v1.11h-1.69v-1.22c0-.75-.31-1.12-.93-1.12s-.93.37-.93,1.12v6.15c0,.74.31,1.11.93,1.11s.93-.37.93-1.11v-1.63h1.69Z'/%3e%3cpath%20class='st0'%20d='M81.4,31.14h-1.8l-.31-2.07h-2.19l-.31,2.07h-1.64l1.82-11.39h2.62l1.82,11.39ZM75.9,19.2l1.52-1.91h1.71l1.51,1.91h-1.61l-.76-.95-.75.95h-1.61ZM77.32,27.52h1.73l-.87-5.76-.87,5.76ZM83.1,15.99l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M84.86,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM84.01,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M93.51,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM92.66,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3cpath%20class='st0'%20d='M98.8,31.14h-1.79v-11.39h1.79v4.88h2.03v-4.88h1.83v11.39h-1.83v-4.88h-2.03v4.88Z'/%3e%3cpath%20class='st0'%20d='M105.36,24.55h2.46v1.62h-2.46v3.34h3.09v1.63h-4.88v-11.39h4.88v1.63h-3.09v3.18ZM108.17,17.29l-1.76,1.91h-1.26l1.17-1.91h1.86Z'/%3e%3cpath%20class='st0'%20d='M112.2,19.75c1.78,0,2.67.94,2.67,2.82v1.48c0,1.87-.89,2.81-2.67,2.81h-.85v4.28h-1.79v-11.39h2.64ZM111.35,21.37v3.86h.85c.58,0,.87-.36.87-1.08v-1.71c0-.71-.29-1.07-.87-1.07h-.85Z'/%3e%3c/g%3e%3ccircle%20class='st1'%20cx='25'%20cy='25'%20r='20'/%3e%3cpath%20class='st0'%20d='M32.78,19.27c2.92,0,4.43,2.55,5.28,5.33l.71,2.17c.14.38-.33.75-.71.75h-5.61c.19-.33.24-.71.09-1.08l-.75-2.45c-.43-1.32-.99-2.64-1.79-3.77.75-.57,1.65-.94,2.78-.94h0ZM25,18.38c3.25,0,4.9,2.78,5.89,5.89l.76,2.45c.14.42-.33.8-.8.8h-11.69c-.42,0-.94-.38-.8-.8l.75-2.45c.99-3.11,2.64-5.89,5.89-5.89h0ZM25,11.35c1.74,0,3.11,1.37,3.11,3.11s-1.37,3.11-3.11,3.11-3.11-1.41-3.11-3.11,1.41-3.11,3.11-3.11h0ZM17.27,19.27c1.08,0,1.98.38,2.73.94-.8,1.13-1.37,2.45-1.74,3.77l-.8,2.45c-.14.38-.05.75.09,1.08h-5.56c-.42,0-.9-.38-.75-.75l.71-2.17c.9-2.78,2.41-5.33,5.33-5.33h0ZM17.27,12.91c1.51,0,2.78,1.27,2.78,2.83s-1.27,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM32.78,12.91c1.56,0,2.78,1.27,2.78,2.83s-1.23,2.83-2.78,2.83-2.83-1.27-2.83-2.83,1.27-2.83,2.83-2.83h0ZM27.07,28.56v.09c0,.57-.24,1.08-.61,1.46h0v.05c-.38.33-.9.57-1.46.57s-1.08-.24-1.46-.61h0c-.38-.38-.61-.9-.61-1.46v-.09h1.41v.09c0,.19.05.38.19.47v.05c.09.09.28.19.47.19s.38-.09.47-.19v-.05c.14-.09.24-.28.24-.47t-.05-.09h1.41ZM30.99,28.56v.09c0,1.65-.66,3.16-1.74,4.24-1.08,1.08-2.59,1.79-4.24,1.79s-3.16-.71-4.24-1.79l-.05-.05c-1.04-1.08-1.7-2.55-1.7-4.2v-.09h1.41v.09c0,1.27.47,2.4,1.27,3.25h.05c.85.85,1.98,1.37,3.25,1.37s2.4-.52,3.25-1.37c.85-.8,1.37-1.98,1.37-3.25v-.09h1.37ZM34.99,28.56v.09c0,2.78-1.13,5.28-2.92,7.07-1.79,1.79-4.29,2.92-7.07,2.92s-5.23-1.13-7.07-2.92c-1.79-1.79-2.92-4.29-2.92-7.07v-.09h1.41v.09c0,2.4.94,4.53,2.5,6.08,1.56,1.56,3.72,2.5,6.08,2.5s4.52-.94,6.08-2.5c1.56-1.56,2.5-3.68,2.5-6.08v-.09h1.41Z'/%3e%3c/svg%3e)