
MINIREVIEW
Osmotic stress sensing and signaling in fishes
Diego F. Fiol and Dietmar Ku
¨ltz
Physiological Genomics Group, Department of Animal Science, University of California, Davis, CA, USA
Physiological significance of osmotic
stress for fishes
Fishes represent the most ancient of five vertebrate
classes. They originated more than 500 million years
ago and have diverged into three major taxa: (a) hag-
fishes and lampreys (Agnatha); (b) cartilagenous fishes
(Chondrichthyii); and (c) ray-finned fishes (Actino-
pterygii). These three taxa employ different strategies
of systemic osmoregulation with only ray-finned fishes
being strong osmoregulators. Nevertheless, at the cellu-
lar level, all fish taxa (like other organisms) ionoregu-
late to maintain K
+
and other intracellular inorganic
ion concentrations within a tightly regulated range,
which is essential to support cell metabolism.
Like other aquatic (or semiaquatic) vertebrates (e.g.
amphibians, alligators), fish are in direct contact with
environmental water. Most fishes depend on stable
water salinity to be able to osmoregulate and maintain
constant osmolality in their body fluids (internal
milieu). These are stenohaline species that can only live
in either freshwater or seawater. Nonetheless, there are
also numerous fish species that tolerate and even thrive
in water characterized by greatly fluctuating salinity.
Keywords
euryhaline fishes; osmoregulation;
osmosensing; osmotic stress; salinity
adaptation; stress signaling
Correspondence
D. Ku
¨ltz, Comparative Physiological
Genomics Group, Department of Animal
Science, One Shields Avenue, Meyer Hall,
University of California, Davis, CA 95616,
USA
Fax: +1 530 752 0175
Tel: +1 530 752 2991
E-mail: dkueltz@ucdavis.edu
(Received 2 July 2007, accepted
7 September 2007)
doi:10.1111/j.1742-4658.2007.06099.x
In their aqueous habitats, fish are exposed to a wide range of osmotic con-
ditions and differ in their abilities to respond adaptively to these variations
in salinity. Fish species that inhabit environments characterized by signifi-
cant salinity fluctuation (intertidal zone, estuaries, salt lakes, etc.) are eury-
haline and able to adapt to osmotic stress. Adaptive and acclimatory
responses of fish to salinity stress are based on efficient mechanisms of
osmosensing and osmotic stress signaling. Multiple osmosensors, including
calcium sensing receptor likely act in concert to convey information about
osmolality changes to downstream signaling and effector mechanisms. The
osmosensory signal transduction network in fishes is complex and includes
calcium, mitogen-activated protein kinase, 14-3-3 and macromolecular
damage activated signaling pathways. This network controls, among other
targets, osmosensitive transcription factors such as tonicity response ele-
ment binding protein and osmotic stress transcription factor 1, which, in
turn, regulate the expression of genes involved in osmotic stress acclima-
tion. In addition to intracellular signaling mechanisms, the systemic
response to osmotic stress in euryhaline fish is coordinated via hormone-
and paracrine factor-mediated extracellular signaling. Overall, current
insight into osmosensing and osmotic stress-induced signal transduction in
fishes is limited. However, euryhaline fish species represent excellent models
for answering critical emerging questions in this field and for elucidating
the underlying molecular mechanisms of osmosensory signal transduction.
Abbreviations
CaSR, calcium sensing receptor; IEG, immediate early gene; MAPK, mitogen-activated protein kinase; Ostf1, osmotic stress transcription
factor 1; TonEBP, tonicity response element binding protein; TRP, transient receptor potential.
5790 FEBS Journal 274 (2007) 5790–5798 ª2007 The Authors Journal compilation ª2007 FEBS
Some of these euryhaline fish (e.g. tilapia) are able to
live in freshwater as well as in water with salinities up
to four times that of seawater. Thus, euryhaline fishes
are able to inhabit environments characterized by
severe osmotic stress, such as desert lakes, tidepools
and estuaries.
Euryhaline fishes have evolved physiological mecha-
nisms that allow them to compensate the osmotic
stress associated with fluctuating environmental salin-
ity. An integral part of such physiological mechanisms
is the ability to sense and quantify changes in environ-
mental salinity and to activate appropriate compensa-
tory responses. Thus, euryhaline fishes represent
excellent models to identify and understand elements
and mechanisms controlling the physiological and
behavioral changes that occur in response to osmotic
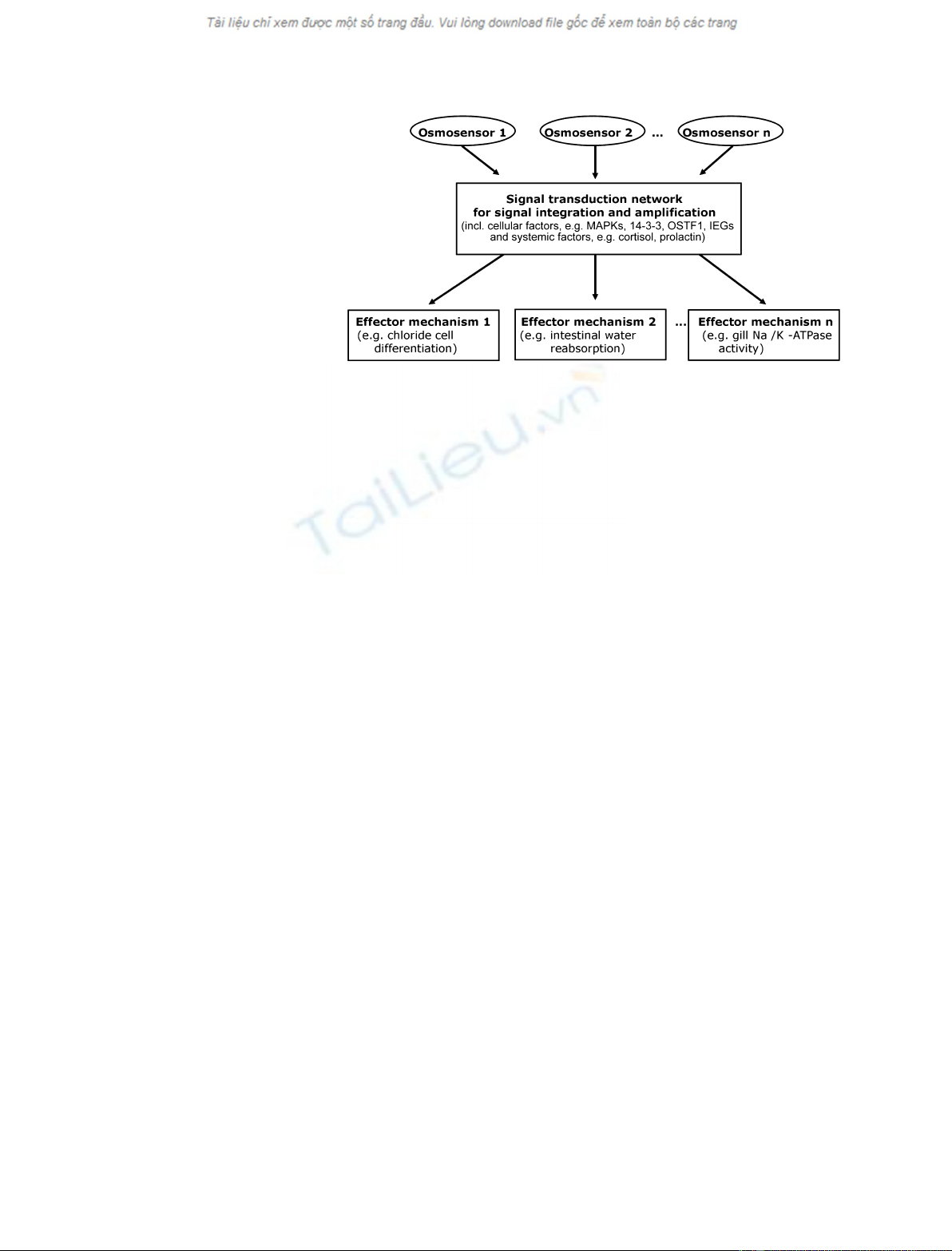
stress. The three major groups of players involved in
this response are osmosensors, signal transducers and
effectors. Osmosensors control signal transduction net-
works that, in turn, regulate effector mechanisms
responsible for acclimation to changes in environmen-
tal salinity (Fig. 1). Many effector mechanisms
involved in osmotic acclimation of euryhaline fishes
have been identified and characterized in detail but
little is known about the proximal osmosensors and
signal transduction pathways that control these effec-
tor mechanisms. In what follows, we will briefly review
our current knowledge about osmosensory signal
transduction in euryhaline fishes and compare it with
knowledge available for some other animals.
Osmotic stress sensing in fishes
General considerations about molecular
osmosensors
Our knowledge of molecular osmosensors that monitor
and quantify environmental and extracellular osmolal-
ity in fishes is minimal. As is true for other cells, it is
not clear how fish cells quantify osmolality to mount a
compensatory adaptive response of proper magnitude
or, alternatively, induce programmed cell death when
their tolerance limit is exceeded. Many different types
of molecular osmosensors can be envisioned because
osmotic stress impacts essentially all cellular structures,
processes and macromolecules. Potential osmosensors
include membrane proteins that are regulated by ion
concentration or membrane stretching and compac-
tion, molecular chaperones that monitor the degree of
protein unfolding, DNA damage sensors, proteins
associated with cytoskeletal organization, and enzymes
whose activity is stringently correlated with intracellu-
lar electrolyte concentration.
It is likely that multiple molecular osmosensors act
in concert to control osmosensory signal transduction
networks and that some of them are activated more
over a range of mild osmotic stress whereas others are
activated more over a range of severe osmotic stress.
In addition, many proximal events perceived by poten-
tial osmosensors (cell volume changes, changes in
cytoskeletal organization, membrane stretching or
compaction, molecular crowding) are only prevalent
during more severe and acute osmotic stress. Such
events result from osmosis across semipermeable mem-
branes of animal cells but osmosis may not occur
when osmolality changes happen gradually over an
extended period of time. Under these conditions, it is
most likely intracellular ionic strength that serves as
the initial signal triggering molecular osmosensors.
Equilibration of intracellular ionic strength during
gradual osmolality changes can be achieved without
osmosis if the capacity of ion transport proteins in cell
membranes is sufficient for moving ions across mem-
branes at a rate that offsets water movement across
membranes. This is only possible for small and slow
osmolality changes and it depends on cell type-specific
Fig. 1. Major elements of the osmosensory
signal transduction network in fishes. Multi-
ple osmosensors (see text) recognize
osmolality ⁄salinity changes and activate a
signaling network that integrates the infor-
mation received from different osmosen-
sors, amplifies this information, and turns
on ⁄off a large number of appropriate effec-
tor mechanisms (i.e. mechanisms of physio-
logical acclimation).
D. F. Fiol and D. Ku
¨ltz Osmotic stress sensing and signaling in fishes
FEBS Journal 274 (2007) 5790–5798 ª2007 The Authors Journal compilation ª2007 FEBS 5791

composition of the cell membrane, including the abun-
dance of particular ion channels, ion transport pro-
teins, aquaporins and membrane lipids. Thus, different
cell types within the same organism may be able to
sense different ranges of osmolality changes. This abil-
ity is critical for euryhaline fishes and other aquatic
vertebrates because some of their cells (e.g. gill cells)
are exposed directly to the aquatic environment and
experience very wide ranges of osmolality whereas
most other cells are bathed in a more homeostatic
environment as a result of systemic osmoregulation.
Changes in extracellular fluid osmolality (i.e. plasma
osmolality) in aquatic vertebrates such as fishes are
also sensed via perception of concomitant fluid volume
and ⁄or blood pressure changes. Systemic osmosensors
and baroreceptors are responsible for monitoring
plasma osmolality and they are conserved in all verte-
brates. Peripheral systemic osmosensors of fishes
appear to be located primarily in the gills [1] and pitui-
tary gland [2]. Once triggered, molecular and systemic
osmosensors activate a signaling network that, in turn,
controls effector mechanisms mediating physiological
acclimation to osmotic stress.
Putative molecular osmosensors in fish cells
Molecular osmosensors of fish cells are not well char-
acterized. However, analysis of zebrafish and pufferfish
genomes shows that putative molecular osmosensors of
mammalian and invertebrate cells are highly conserved
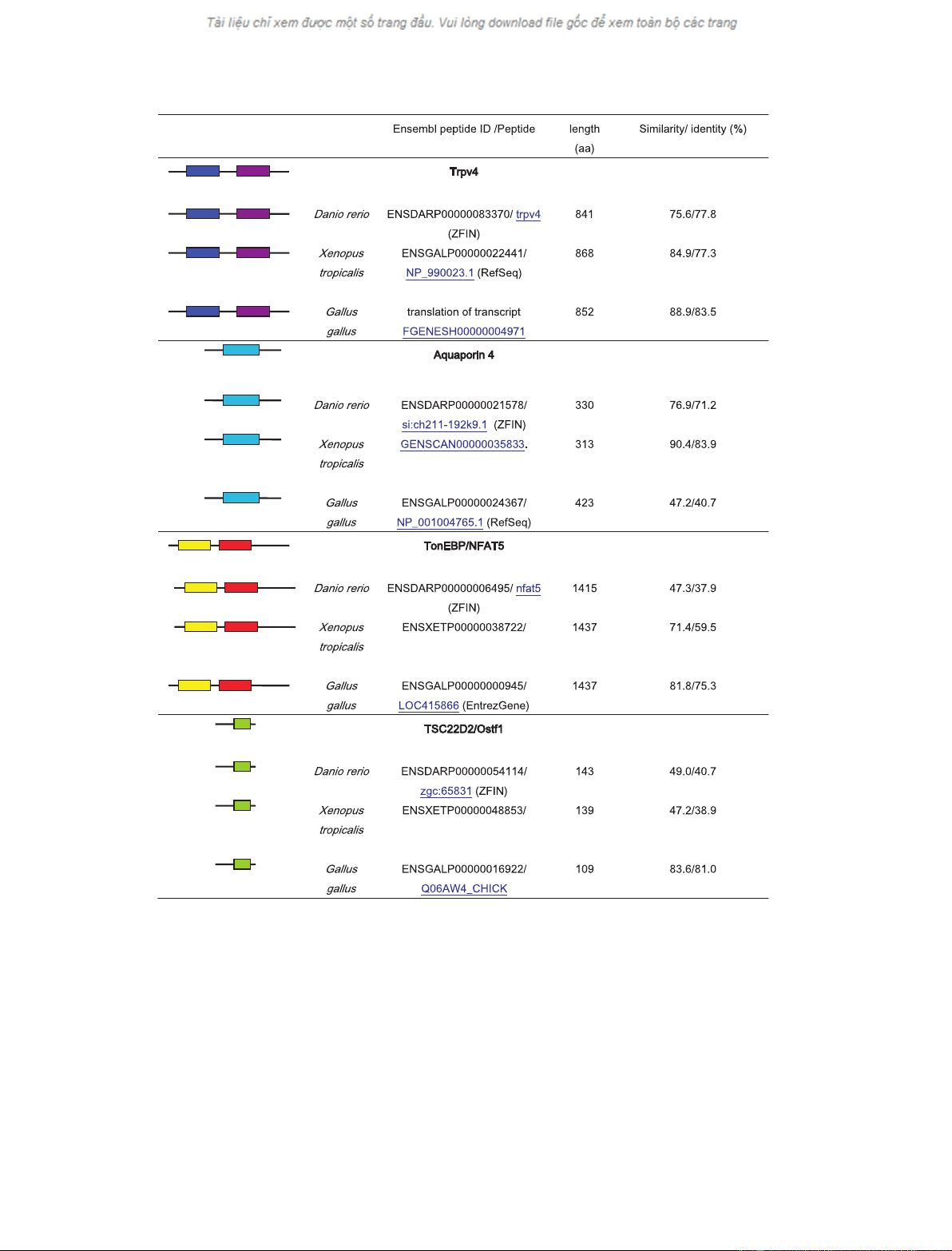
in fish genomes (Fig. 2). Such putative osmosensors
include adenyl cyclase [3], transient receptor potential
(TRP) channels [4], and aquaporin 4 [5]. However,
functional evidence firmly establishing these proteins
as molecular osmosensors in euryhaline fishes is lack-
ing and this area needs to be experimentally addressed
in future research.
A role of adenyl cyclase as an osmosensor in eury-
haline fishes is supported by its effects on chloride
secretion across the gill epithelium and osmoregulatory
hormone secretion from the pituitary gland. Forskolin,
which stimulates adenyl cyclase activity, was shown to
enhance chloride secretion across opercular membranes
of euryhaline fishes [6], as well as prolactin and growth
hormone secretion from trout pituitary gland [7].
These secretory processes are also stimulated when
euryhaline fish face salinity increases. Nevertheless, it
is not known whether activation of adenyl cyclase in
euryhaline fishes is directly mediated by osmolality
changes as would be required for a true osmosensor
protein.
Osmosensory TRPV4 channels were localized in
Danio rerio and the expression of this channel protein
in the developing kidney was demonstrated [8]. More-
over, evolutionary studies on the TRP protein family
identified six copies of TRPV4 in the western clawed
frog (Xenopus tropicalis), suggesting that diversification
of osmosensory TRPV4 may favor adaptation to both
aquatic and terrestrial environments, which represent
very different habitats regarding requirements for
osmoregulation [9]. However, as for adenyl cyclase, no
direct evidence for an osmosensory function of TRP
channels in fishes has been published.
Aquaporin water channels have been studied in
fishes, including their regulation during salinity stress.
Nevertheless, all studies to date have focused on the
role of aquaporins as effector proteins of osmosensory
signal transduction pathways and the potential role of
these proteins in osmosensing of fish cells has yet to
be addressed. As a result of recent studies on fish aqu-
aporins, we know that changes in water permeability
in gills and intestine are mediated at least in part via
regulation of aquaporin abundance in epithelial cell
membranes [10]. Thus, a role of aquaporins as impor-
tant effector proteins of osmoregulation in fishes has
been established. It will be interesting to see whether
water channels also function as systemic osmosensors
in the brain of fishes, as has been suggested for
mammals [5].
Calcium sensing receptor
The calcium sensing receptor (CaSR) has been identi-
fied as an important osmosensor protein in fishes.
CaSR is a large glycoprotein belonging to the G pro-
tein-coupled receptor superfamily. This membrane pro-
tein is regulated directly by extracellular calcium (and
to some extent also other polyvalent cations) as ligand
in the millimolar range. Fishes (e.g. euryhaline marine
species) utilize CaSR for sensing environmental salinity
[11]. In particular for marine fishes, the calcium con-
centration in the external environment (seawater) is in
the millimolar range that is accurately sensed by
CaSR. Thus, changes in environmental calcium con-
centration are thought to be a surrogate measure for
the ionic strength ⁄salinity of the marine environment.
In agreement with this notion, CaSR is expressed in
osmoregulatory tissues of fishes, including shark rectal
gland [12] and teleost gill and opercular membrane
[13]. Full-length transcripts of CaSR have been cloned
from gilthead sea bream (Sparus aurata) [13] and spiny
dogfish (Squalus acanthias) [11]. Using nucleotide
probes, CaSR transcripts have been localized to bran-
chial chloride cells of both aforementioned species, as
well as winter flounder (Pleuronectes americanus) and
Atlantic salmon (Salmo salar) [11].
Osmotic stress sensing and signaling in fishes D. F. Fiol and D. Ku
¨ltz
5792 FEBS Journal 274 (2007) 5790–5798 ª2007 The Authors Journal compilation ª2007 FEBS
Tilapia CaSR senses changes in external [Ca
2+
] and
activates phospholipase C and mitogen-activated pro-
tein kinase (MAPK) signaling [14]. Moreover, changes
in plasma [Ca
2+
] and [Mg
2+
] that occur when fish
move from freshwater to seawater, or vice versa, likely
serve as salinity sensing cues for CaSR because plasma
4e-19 5e-32
2e-18 4e-35
9e-18 3e-35
RHD IPT_NFAT
1e-08
2e-15
3e-15
TSC22
1e-53
3e-45
1e-52
M/P
3e-10 4e-10
1e-12 7e-10
3e-12 4e-10
ANK ION_TRANS
Fig. 2. Evolutionary conservation of orthologs of the putative osmosensors TRPV4 and aquaporin 4 and the osmosensory signal transcription
proteins TonEBP and TSC22D2 in vertebrates. The human sequences of TRPV4 (871 amino acids, AAG28029.1), aquaporin 4 (323 amino
acids, NP_001641.1), TonEBP (1531 amino acids, NP_006590.1), and the mouse sequence of TSC22D2-4 (116 amino acids, EU004151) were
used as references and their conserved domains are indicated. The highest homology hits for each D. rerio,X. tropicalis and Gallus gallus ge-
nomes were analyzed for the presence of conserved domains in the Conserved Domain Database and Search Service, version 2.11 (NCBI,
17402 motifs) and the expectation values are indicated. Percentages of amino acid sequence similarity and identity are shown. RHD, Rel
homology domain (pfam00554); IPT_NFAT, IPT domain of the NFAT family of transcription factors (cd01178); TSC22, TSC-22 ⁄dip ⁄bun family
(pfam01166); MIP, major intrinsic protein (cd00333); ANK, ankyrin repeats (cd00204); ION_TRANS, ion transport protein (pfam00520).
D. F. Fiol and D. Ku
¨ltz Osmotic stress sensing and signaling in fishes
FEBS Journal 274 (2007) 5790–5798 ª2007 The Authors Journal compilation ª2007 FEBS 5793

concentrations of these divalent ions are well within
the millimolar range over which CaSR operates [11].
In the mammalian kidney, CaSR regulates the activity
of many other signaling pathways, including pathways
that are regulated by intracellular calcium concentra-
tion. The evidence briefly summarized above suggests
that CaSR plays a significant role for osmosensing in
fishes.
Osmosensory signal transduction
network in fishes
Studies on bacteria, yeast and model animals have
shown that osmosensors control an elaborate intracel-
lular signaling network. The major role of this network
is to integrate signals from multiple osmosensors and
generate an amplified output-stimulus for controlling
appropriate effector mechanisms (Fig. 1). We hypothe-
size that the mode of integration of signals generated
by multiple osmosensors with different sensitivity
ranges enables cells to determine the severity of osmo-
tic stress, quantify extracellular osmolality, and ensure
that an appropriate physiological response is mounted.
Testing this hypothesis will require detailed knowledge
about the key elements involved in osmosensory signal
transduction. Known elements of osmosensory signal
transduction in euryhaline fishes are calcium-dependent
pathways, MAPKs, 14-3-3 proteins, specific transcrip-
tion factors, hormones, and paracrine factors. Their
role during osmotic stress is briefly reviewed below.
Role of intracellular calcium
We have summarized above that environmental cal-
cium may be an important trigger of osmosensory
events by controlling CaSR activity. In addition, many
effects of changes in environmental and plasma cal-
cium concentration on fish gill chloride cell morphol-
ogy and the function of important osmoregulatory
effector proteins have been documented [15]. Since cal-
cium is a major second messenger in eukaryotic cells
and known to play significant roles in osmosensory
signal transduction of mammalian and even plant cells,
it is very likely that calcium-mediated signaling con-
tributes significantly to osmosensory signal transduc-
tion in fish cells. The importance of intracellular
calcium for the activation of downstream signaling
events in fish exposed to osmotic stress has been stud-
ied in fish rostral pars distalis cells. These cells are
excellent models because they represent a relatively
homogeneous (approximately 97%) population of pro-
lactin secreting cells and their prolactin secretion
depends on osmolality. In tilapia, hyposmotic stress
stimulates prolactin secretion, which was shown to
depend on stretch-activated ion channels and increased
intracellular calcium [16]. Cortisol, a hormone associ-
ated with hyperosmotic stress, inhibits prolactin secre-
tion via reduction of free intracellular calcium [17]. In
addition to its effect on intracellular calcium, cortisol
also inhibits adenyl cyclase, a potential osmosensor
mentioned above, suggesting that both major intra-
cellular second messengers, calcium and cAMP, are
involved in osmotic stress signaling [18]. Another
osmoregulatory hormone, angiotensin II, increases free
intracellular calcium in fish tissues [19], confirming that
the effects of osmoregulatory hormones are mediated
at least partly via intracellular calcium signaling. An
important role of intracellular calcium in fish osmotic
stress signaling is also supported by a modeling
approach yielding an osmosensory signal transduction
network based on 20 immediate early genes that rap-
idly respond to salinity stress in tilapia gill. Intracellu-
lar calcium is a major node in this network, which also
contains several calcium-binding proteins such as an-
nexins and S-100 proteins [20]. Notably, annexins and
two other immediate early genes (IEGs) identified in
this study (gelsolin, galectin 4) are known to regulate
actin-based cytoskeleton remodeling in mammalian
cells, suggesting that this process may be a major tar-
get during osmotic stress acclimation in fish gill cells.
Consistent with this view, the actin-based cytoskeleton
seems to play a role in osmotic regulation of
Na
+
⁄K
+
⁄2Cl
–
(NKCC) cotransporter [21] and in the
closing or opening of apical crypts of gill chloride cells
[22]. Furthermore, changes in ion transport during
hyper- and hypotonic stress require intact F-actin and
microtubules in eel intestinal epithelium [23].
MAPK
MAPKs are a family of enzymes that are involved in
osmosensory signal transduction in yeast, plant and
animal cells. They are key elements of protein phos-
phorylation cascades that integrate and amplify signals
from osmosensors to activate appropriate downstream
targets mediating physiological acclimation. Although
MAPKs are highly evolutionarily conserved, their acti-
vators and substrates can differ greatly, depending on
taxon, physiological condition and developmental
state. For example, yeast exposed to osmotic stress
activate the high osmolarity glycerol response (HOG1)
MAPK cascade via the SLN1 osmosensor, which is a
two-component histidine kinase, none of whose com-
ponents are present in any sequenced animal genome.
This illustrates that osmotic stress signaling networks
are modular. Recent evidence suggests that MAPK
Osmotic stress sensing and signaling in fishes D. F. Fiol and D. Ku
¨ltz
5794 FEBS Journal 274 (2007) 5790–5798 ª2007 The Authors Journal compilation ª2007 FEBS

























