Chương 7
CƠ SỞ HÓA LÝ CỦA SỰ HƯNG PHẤN
I. Khái niệm hưng phấn và ngưỡng hưng phấn
* Khái niệm hưng phấn
Hưng phấn là sự chuyển từ trạng thái nghỉ ngơi sang trạng thái hoạt động. Hưng phấn bao
gồm hai cơ chế: Cơ chế tiếp nhận kích thích bởi các thụ quan và cơ chế chuyển tín hiệu
kích thích thành tín hiệu điện, truyền về não để xử lý thông tin và phát tín hiệu thực hiện
phản ứng trả lời. Tín hiệu kích thích rất đa dạng nhưng chủ yếu là tín hiệu vật lý (nhiệt, ánh
sáng, áp suất...) và tín hiệu hóa học (hoócmôn, mùi, vị...). Chức năng chuyển tín hiệu kích
thích thành tín hiệu điện (tức sóng hưng phấn) và dẫn truyền sóng hưng phấn do noron thực
hiện. Thực hiện phản ứng trả lời có thể là cơ quan, mô, tế bào và cả ở mức độ phân tử.
Trong hệ sinh vật, từ sinh vật đơn bào tới sinh vật đa bào tuy có mức độ tiến hóa khác xa
nhau nhưng đều tồn tại tính hưng phấn để thích nghi với sự thay đổi của môi trường sống.
* Khái niệm ngưỡng hưng phấn
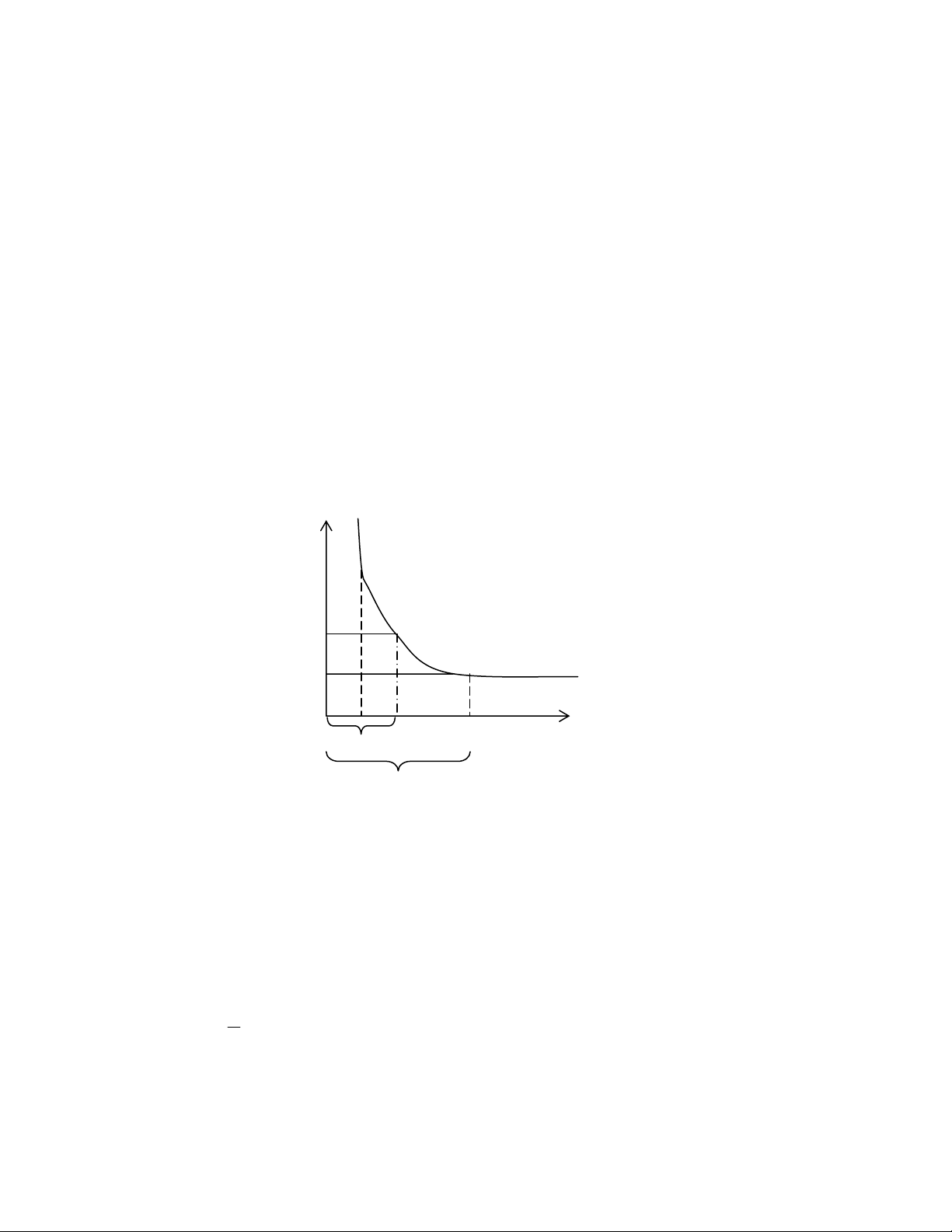
E
D
B
C
O
1 Reobaz A
2 Reobaz G
Cư
ờ
n
g
đ
ộ
Thời gian
Thời trị
Thời
g
ian có ích
Hình 7.1: Tương quan giữa cường độ và thời gian kích thích
Ngưỡng hưng phấn được xác định bằng cường độ nhỏ nhất và thời gian kích thích ngắn
nhất để có thể tạo nên sự hưng phấn. Cường độ nhỏ nhất kích thích để tạo ra được phản
ứng trả lời gọi là 1 reobaz. Thời gian ngắn nhất khi kích thích 1 reobaz để tạo ra được
phản ứng trả lời là thời gian có ích (xem hình 7.1). Trong thực nghiệm xác định thời gian
có ích rất khó nên Lapicque lấy thời gian ứng với 2 reobaz để đo ngưỡng thời gian kích
thích, gọi là thời trị. Đường biểu diễn tương quan giữa cường độ và thời gian kích thích là
đường hipecbol, ứng với phương trình do Weiss đưa ra năm 1901:
b
t
a
i+= (7.1)
i: Cường độ ngưỡng
t: Thời gian ngưỡng

a: Hằng số ứng với đường thẳng thời gian chạy song song với trục tung
b: Hằng số ứng với đường thẳng cường độ chạy song song với trục hoành
Nếu cường độ i = 2b, nghĩa là bằng 2 reobaz thì phương trình (7.1) sẽ có dạng:
2b = t
a + b (7.2) → b =
t
a → t =
b
a
Thời trị thay đổi tùy theo mô. Ví dụ ở người thời trị của cơ duỗi dài gấp từ 1,5 đến 2 lần
so với cơ gập.
II. Lý thuyết hưng phấn của Heinbrun (1928)
Trên cơ sở những số liệu thực nghiệm về quá trình hưng phấn có liên quan tới sự thay
đổi cấu trúc hóa lý của nguyên sinh chất, như là thay đổi tính chất keo thể hiện ở tế bào
thực vật bậc cao và động vật nguyên sinh (amip) mà Heinbrun đã đưa ra thuyết đông tụ
vào năm 1928. Heinbrun cho rằng: Tất cả các yếu tố kích thích đều gây nên quá trình
đông tụ nguyên sinh chất kèm theo sự tăng đột ngột độ nhớt cấu trúc của nó. Quan niệm
này được củng cố qua số liệu về các chất ức chế sự hưng phấn như thuốc ngủ, thuốc mê
đều làm giảm độ nhớt của nguyên sinh chất.
Thuyết đông tụ của Heinbrun giải thích các yếu tố kích thích có bản chất khác nhau khi
tác dụng lên các tế bào của thụ quan, trước tiên giải phóng Ca++ mà trước đó trong
nguyên sinh chất Ca++ lại ở trạng thái liên kết. Chính do Ca++ được giải phóng đã dẫn tới
làm đông tụ nguyên sinh chất. Thuyết đông tụ của Heinbrun mới chỉ giải thích được hiện
tượng sự kích thích dẫn tới làm đông tụ nguyên sinh chất còn nhiều hiện tượng khác liên
quan tới sự hưng phấn, thuyết đông tụ không giải thích được.
III. Thuyết phá hủy cấu trúc của Naxonov và Alecxandrov (1940-1943)
Naxonov và Alecxandrov đã xem quá trình hưng phấn như là một quá trình phá hủy cấu
trúc. Hai ông quan niệm nguyên sinh chất là pha không hòa tan trong nước và sự phân bố
không đồng đều của các chất ở trong nội bào và ngoài môi trường là do khả năng hòa tan
của các chất ở pha nước và nguyên sinh chất khác nhau và do khả năng liên kết của các
chất với phân tử protein. Trong tế bào các chất điện phân phần lớn liên kết với các phân
tử protein, chỉ số ít ở trạng thái tự do. Khi bị kích thích hay bị tổn thương nguyên sinh
chất có những thay đổi sau:
- Độ phân tán của các hạt keo giảm xuống rõ rệt và độ đục của hạt nhân và nguyên sinh
chất xuất hiện rất sớm.
- Sự tăng độ nhớt của nguyên sinh chất gồm 2 pha: Khi yếu tố kích thích yếu thì độ nhớt
nguyên sinh chất giảm và khi yếu tố kích thích tăng lên thì độ nhớt nguyên sinh chất tăng
lên rất nhanh.
- Khi kích thích, ban đầu quá trình tạo hạt trong nguyên sinh chất tăng lên và sau đó quá
trình này bị ức chế.
- Khi kích thích độ pH của nguyên sinh chất dịch chuyển về phía axit.
- Khi kích thích K+, , creatin được giải phóng ra môi trường còn Na
−3
4
PO +, Cl- lại xâm
nhập vào trong tế bào rất nhanh.

Cũng như thuyết đông tụ, thuyết phá hủy cấu trúc đưa ra chỉ là để giải thích về sự thay
đổi tính chất hóa lý của nguyên sinh chất, mới giải thích hiện tượng của quá trình hưng
phấn mà chưa giải thích được bản chất của quá trình hưng phấn.
IV. Lý thuyết hưng phấn của Nernst (1899)
Năm 1887, Arenius công bố các chất khi hòa tan trong nước sẽ phân ly thành các ion
dương và ion âm, dưới tác dụng của dòng điện ngoài, các ion sẽ chạy về điện cực mang
điện tích trái dấu với điện tích ion. Các ion có kích thước và bản chất điện tích khác nhau
nên có vận tốc chuyển động về hai cực cũng khác nhau. Sau khoảng thời gian nhất định
sẽ tạo nên những lớp có mật độ ion khác nhau, tức sẽ xuất hiện một hiệu điện thế. Nếu
ngắt nguồn điện bên ngoài, thay vào đó cắm 2 điện cực vào dung dịch điện phân, nối với
một bóng điện thì đèn sẽ sáng. Đó là dòng điện xuất hiện trong dung dịch điện phân. Năm
1899, Nernst dựa trên kết quả nghiên cứu của Arenius cũng xem tế bào như một dung
dịch chất điện phân được bao bọc bởi màng tế bào. Dưới tác dụng của dòng điện kích
thích, các ion âm và dương trong tế bào chất sẽ chạy về hướng điện cực kích thích có
điện tích trái dấu với điện tích ion. Sau một thời gian các ion âm và dương chuyển động
theo hai hướng khác nhau sẽ tập trung ở hai phía của màng tế bào. Ở ngoại bào cũng có
các ion dương và âm, do lực hút tĩnh điện, nếu ở một phía tế bào, mặt trong tích điện âm
thì mặt ngoài màng tích điện dương còn ở phía kia của tế bào ở mặt trong sẽ tích điện
dương và mặt ngoài màng tích điện âm. Kết quả là giữa bên trong và bên ngoài màng tế
bào đã hình thành nên một hiệu điện thế và khi điện thế này đạt giá trị ngưỡng thì tạo ra
sự hưng phấn. Sự chênh lệch về nồng độ ion liên quan đến cường độ và thời gian kích
thích và để tạo ra hưng phấn phải thỏa mãn công thức:
k
t
.i.CC oγ=− (7.3)
C: Nồng độ ion tự do khi tế bào hưng phấn
Co: Nồng độ ion tự do khi tế bào ở trạng thái nghỉ ngơi
γ: Số lượng ion được dịch chuyển do một đơn vị cường độ dòng điện
i: Cường độ dòng điện; t: Thời gian kích thích
k: Hệ số khuyếch tán của ion
Nếu kích thích bằng dòng điện một chiều thì mối liên quan giữa cường độ dòng điện và
thời gian kích thích phải thỏa mãn công thức:
ti = hằng số (7.4)
Trong giới hạn về cường độ dòng điện kích thích thì nếu cường độ dòng điện tăng thì
thời gian kích thích giảm và ngược lại, để duy trì tích số của cường độ dòng điện với thời
gian luôn là một hằng số.
Nếu kích thích bằng dòng điện xoay chiều thì mối liên quan giữa cường độ và tần số
dòng điện phải thỏa mãn công thức:
ω
i= hằng số (7.5)
Trong giới hạn về cường độ dòng điện kích thích, nếu tần số cao thì cường độ dòng điện
phải lớn còn tần số thấp thì cường độ dòng điện nhỏ, để duy trì tỷ số giữa cường độ và

tần số luôn là một hằng số. Các công thức trên do Nernst đưa ra chỉ đúng trong phạm vi
tần số 100 Hz đến 3 kHz.
Kết quả thực nghiệm cho thấy nếu kích thích cường độ dưới ngưỡng và kích thích lâu
hoặc kích thích cường độ lớn với thời gian kích thích ngắn thì đều không gây ra hưng
phấn. Ở mỗi đối tượng nghiên cứu khi kích thích cường độ dòng điện là 1 reobaz thì có
một thời gian kích thích cần thiết (thời gian hữu ích).
Bảng 7.1: Thời gian hữu ích của một số đối tượng nghiên cứu
TT
Đối tượng nghiên cứu Thời gian hữu ích
1 Cơ trơn dạ dày ếch 1 giây
2 Cơ trơn của nhuyễn thể 10-1 giây
3 Cơ chân của nhuyễn thể 10-2 giây
4 Thần kinh ếch 10-3 giây
5 Thần kinh động vật máu nóng 10-4 giây
Lý thuyết hưng phấn của Nernst không nêu ra cụ thể sự thay đổi nồng độ của những ion
nào và giới hạn ngưỡng nồng độ là bao nhiêu để có thể tạo ra hưng phấn.
V. Lý thuyết hưng phấn của Bernstein (1906)
Năm 1906, Bernstein đưa ra lý thuyết hưng phấn để giải thích cơ chế hình thành điện
thế tĩnh khi tế bào ở trạng thái nghỉ ngơi và cơ chế hình thành điện thế hoạt động khi tế
bào ở trạng thái hưng phấn. Bernstein cho rằng tế bào khi ở trạng thái nghỉ ngơi do màng
có tính bán thấm tức là thấm K+ dễ dàng còn Na+ thấm ít, trong khi đó hoàn toàn không
thấm các phân tử hữu cơ mang điện tích âm (gọi là các amion hữu cơ). Do vậy, tế bào ở
trạng thái tĩnh, bên trong có điện tích âm còn ngoài màng có điện tích dương nên tồn tại
điện thế tĩnh và chiều điện trường hướng từ ngoài vào trong tế bào. Bernstein đơn thuần
chỉ xét sự chênh lệch nồng độ K+ ở bên trong và bên ngoài tế bào, áp dụng công thức của
Nernst tính được giá trị điện thế tĩnh phù hợp với giá trị đo trực tiếp bằng phương pháp vi
điện cực. Sau này có nhiều số liệu thực nghiệm làm sáng tỏ thêm lý thuyết của Bernstein.
Xegan đã chứng minh các ion hóa trị hai (như Ca++, Mg++,...) hút phân tử nước (H2O)
mạnh hơn so với ion hóa trị một (như Na+, K+, Cl-...). Vì thế lớp vỏ hidrat hóa của ion
hóa trị hai dày hơn so với ion hóa trị một, nên ion hóa trị hai khó lọt qua siêu lỗ ở trên
màng so với ion hóa trị một. Mặt khác, cùng một loại ion, nếu ion nào có kích thước nhỏ
(hay nguyên tử lượng bé) sẽ có điện trường lớn so với ion có kích thước lớn (hay nguyên
tử lượng lớn). Vì thế ion có kích thước bé hút nước mạnh hơn so với ion có kích thước
lớn nên xảy ra hiện tượng, trong dung dịch ion có kích nhỏ lại khó thấm qua siêu lỗ trên
màng so với ion có kích thước lớn hơn. Ví dụ Na+ có đường kính 1,9A0 bị 8 phân tử H2O
bao quanh còn K+ có đường kính 2,6A0 chỉ có 4 phân tử H2O bao quanh nên K+ thấm qua
siêu lỗ trên màng dễ hơn so với Na+. Năm 1955, Hodgkin và Katz bằng phương pháp
đồng vị phóng xạ đánh dấu xác định được khi tế bào ở trạng thái tĩnh, dòng Na+ đi ra
bằng dòng Na+ đi vào còn dòng K+ đi ra lớn gấp xấp xỉ 1,63 lần so với dòng K+ đi vào.
Hơn nữa khi tế bào ở trạng thái tĩnh, nồng độ Na+ ở bên ngoài lớn hơn so với bên trong
nên chiều gradien nồng độ Na+ hướng từ ngoài vào trong còn nồng độ K+ bên trong lại
cao hơn bên ngoài nên chiều gradien nồng độ K+ lại hướng từ trong ra ngoài . Song do
tính bán thấm của màng, theo tính toán của Goldman cho thấy K+ đi ra theo gradien
nồng độ K+ nhanh gấp 76 lần so với Na+ đi vào cùng chiều gradien nồng độ Na+. Kết quả

thực nghiệm khẳng định tính đúng đắn của giả thuyết Bernstein, tính bán thấm của màng
là nguyên nhân dẫn đến sự phân bố không đồng đều của các ion giữa bên trong và bên
ngoài màng tế bào nên đã tạo thành điện thế tĩnh. Sau này, năm 1942, Cuatite và Cole lại
chứng minh rằng K+ đóng vai trò chính trong việc duy trì điện thế tĩnh. Hai ông đã tăng
nồng độ K+ ở ngoại bào lên (tức làm giảm sự chênh lệch nồng độ K+ giữa bên trong và
bên ngoài màng tế bào) thì giá trị điện thế tĩnh cũng giảm dần. Khi nồng độ K+ ở bên
ngoài bằng nồng độ K+ ở bên trong tế bào thì điện thế tĩnh sẽ bằng không. Khi giảm dần
nồng độ K+ ở bên ngoài về giá trị như lúc ban đầu thì điện thế tĩnh lại được khôi phục.
Song nếu tăng nồng độ Na+ ở trong tế bào lên thì điện thế tĩnh vẫn không thay đổi. Khi bị
kích thích, tế bào đã chuyển từ trạng thái tĩnh sang trạng thái hưng phấn và tế bào bị đổi
cực, tức là bên trong tế bào có điện tích dương còn bên ngoài có điện tích âm. Khi tế bào
hưng phấn, Bernstein cho rằng màng tế bào đã mất tính bán thấm nên không còn phân
biệt tính thấm giữa K+ với Na+ và cho cả các anion hữu cơ thấm ra bên ngoài. Chính do
các anion hữu cơ thấm ra ngoài và các Na+ thấm vào trong dễ dàng hơn so với K+ thấm ra
ngoài đã làm cho tế bào bị đổi cực. Sau này thực nghiệm đo được giá trị tuyệt đối của
điện thế hoạt động lớn hơn giá trị tuyệt đối của điện thế tĩnh nhưng theo giả thuyết của
Bernstein thì không giải thích được bản chất của hiện tượng này. Ví dụ noron thần kinh
lúc nghỉ ngơi có điện thế tĩnh là -90mV còn khi hưng phấn có điện thế hoạt động từ 120
đến 130mV. Năm 1949, Hodgkin và Katz đã làm thí nghiệm chứng minh rằng khi noron
hưng phấn tính thấm của màng đối với Na+ vào trong tế bào tăng lên 500 lần so với khi
noron nghỉ ngơi. Ngược lại, khi noron nghỉ ngơi màng tế bào cho K+ thấm ra ngoài lại
nhanh gấp 76 lần so với Na+ thấm vào trong tế bào.
Sau này Goldman đưa ra công thức để tính điện thế tĩnh và điện thế hoạt động. Mỗi ion
có một hệ số thấm đặc trưng. Khi noron nghỉ ngơi, nếu lấy tính thấm của màng đối với
K+ làm đơn vị so sánh và qui ước PK+ =1 thì tính thấm của màng đối với Na+ là PNa+ =
0,013 còn tính thấm của màng đối với Cl- là PCl-_= 0,045. Cùng với việc xác định nồng độ
K+, Na+, Cl- ở bên ngoài và bên trong noron lúc nghỉ ngơi, thay các giá trị đã biết vào
công thức Gondman sẽ tính được điện thế tĩnh bằng -89mV. Khi noron hưng phấn nếu
vẫn lấy PK+ = 1 làm đơn vị để so sánh thì PNa+ = 20, PCl- = 0,045. Cũng xác định nồng độ
K+, Na+, Cl- ở bên trong và bên ngoài noron lúc hưng phấn, thay các giá trị đã biết vào
công thức Goldman sẽ tính được điện thế hoạt động bằng 38mV. Khi noron hưng phấn,
điện màng đã chuyển từ -89mV lên 38mV, do vậy giá trị tuyệt đối của điện thế hoạt động
phải là |-89mV|+38mV=127mV. Như vậy, bản chất của hiện tượng điện thế hoạt động có
giá trị tuyệt đối lớn hơn giá trị tuyệt đối của điện thế tĩnh. Khi ngừng kích thích, màng
noron sẽ khôi phục lại tính bán thấm để duy trì sự chênh lệch về nồng độ ion giống như
lúc ban đầu, tức là duy trì điện thế tĩnh.
Lý thuyết màng của Bernstein chưa đề cập đến vai trò của các ion hóa trị hai như Ca++,
Mg++... Nhiều kết quả thực nghiệm đã xác định vai trò của Ca++ trong quá trình hình
thành điện thế sinh vật. Một số nhà khoa học giả thiết rằng, ở trên màng tế bào có các
kênh dẫn "nhanh" và "chậm" đối với các ion. Khi tế bào hưng phấn, các kênh dẫn
"nhanh" cho dòng Na+ vào trong tế bào để khử cực tế bào (tức đổi dấu điện tích âm sang
dương). Sau đó các kênh dẫn "chậm" tiếp tục cho Na+ và Ca++ vào trong tế bào để kết
thúc quá trình khử cực (tức là xuất hiện điện thế hoạt động).
Lý thuyết màng của Bernstein, mặc dù đã được các nhà khoa học bổ sung nhưng vẫn
chưa đầy đủ. Do vậy, vấn đề này vẫn còn phải tiếp tục nghiên cứu và hoàn thiện.
VI. Lý thuyết hưng phấn của Laxarev



![Bài giảng Sinh học đại cương: Sinh thái học [mới nhất]](https://cdn.tailieu.vn/images/document/thumbnail/2025/20250812/oursky02/135x160/99371768295754.jpg)





















